Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2020; 29(3): 219-229
Published online June 30, 2020
https://doi.org/10.5607/en20006
© The Korean Society for Brain and Neural Sciences
Labeling Dual Presynaptic Inputs using cFork Anterograde Tracing System
Jun-Young Oh1†, Jeong-Ho Han2†, Hyoeun Lee2, Young-Eun Han1, Jong Cheol Rah1,3 and Hyungju Park1,2,3*
1Multi-institutional Collaborative Research Center for Cortical Processing, Korea Brain Research Institute (KBRI), Daegu 41062, 2Molecular Neurobiology Lab, Research Group for Neurovascular Unit, Korea Brain Research Institute (KBRI), Daegu 41062, 3Department of Brain and Cognitive Sciences, DGIST, Daegu 42988, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-53-980-8450, FAX: 82-53-980-8339
e-mail: phj2@kbri.re.kr
†These authors contributed equally to this work.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
Understanding brain function-related neural circuit connectivity is essential for investigating how cognitive functions are decoded in neural circuits. Trans-synaptic viral vectors are useful for identifying neural synaptic connectivity because of their ability to be transferred from transduced cells to synaptically connected cells. However, concurrent labeling of multisynaptic inputs to postsynaptic neurons is impossible with currently available trans-synaptic viral vectors. Here, we report a neural circuit tracing system that can simultaneously label postsynaptic neurons with two different markers, the expression of which is defined by presynaptic input connectivity. This system, called “cFork (see fork)”, includes delivering serotype 1-packaged AAV vectors (AAV1s) containing Cre or flippase recombinase (FlpO) into two different presynaptic brain areas, and AAV5 with a dual gene expression cassette in postsynaptic neurons. Our in vitro and in vivo tests showed that selective expression of two different fluorescence proteins, EGFP and mScarlet, in postsynaptic neurons could be achieved by AAV1-mediated anterograde trans-synaptic transfer of Cre or FlpO constructs. When this tracing system was applied to the somatosensory barrel field cortex (S1BF) or striatum innervated by multiple presynaptic inputs, postsynaptic neurons defined by presynaptic inputs were simultaneously labeled with EGFP or mScarlet. Our new anterograde tracing tool may be useful for elucidating the complex multisynaptic connectivity of postsynaptic neurons regulating diverse brain functions.
Graphical Abstract
Keywords: Adeno-associated virus -1, Adeno-associated virus -5, Brain mapping, EGFP, mScarlet, Anterograde tracing
INTRODUCTION
Cognitive functions are regulated by multiple neuronal activities mediated by highly elaborate and complex synaptic connections. Postsynaptic neurons receiving multiple presynaptic inputs process complex presynaptic input activities, and then produce unique activity patterns encoding certain brain functions. Thus, it is necessary to understand what presynaptic inputs are connected to certain postsynaptic neurons. Neuroanatomical tracing molecules that can be transmitted in an anterograde direction have been widely utilized for this purpose.
Neuroanatomical tracers, including biotinylated dextran amines (BDA), horseradish peroxidase (HRP), wheat germ agglutinin (WGA), and cholera toxin B subunit (CTB) [1, 2], can be easily used in animal brains to efficiently label synaptic connectivity. However, these non-viral neuroanatomical tracers are unsuitable for revealing fine neuronal connectivity or the cell types that make synapses. Classical anterograde tracing with viral vectors uses the density of axonal projections as proxies of potential synaptic connections to postsynaptic targets. However, this method cannot distinguish the trespassing axonal collaterals from actual synaptic connections without additional high-resolution imaging. Moreover, the connectivity can be misinterpreted when synaptic connections are formed in a distant dendritic location.
Recent advances using viral vectors have been shown to overcome such limitations, given their innate capability of being transferred from infected cells to other cells via synapses. Viral vectors including adeno-associated virus (AAV), vesicular stomatitis virus (VSV), and herpes simplex virus (HSV), are known to spread in the anterograde direction across synapses and have been proven to express transgenes in the viral genome [2,-7]. However, labeling multiple connectivity in the postsynaptic neuron is limited because only a single population of presynaptic neurons infected by anterograde trans-synaptic viruses is delivered to postsynaptic partner neurons. Since animal cognitive functions and behaviors depend on the complex involvement of multiple neurons and their synaptic crosstalk, the demand for trans-synaptic viral vectors suitable for labeling multiple synaptic connectivity systems has been increasing. It is thus necessary to develop anterograde trans-synaptic viral vectors capable of expressing multiple marker genes, each of which represent specific presynaptic inputs, in the postsynaptic neurons.
Here, we report a dual-color anterograde tracing vector system (cFork) using presynaptic AAV1-mediated delivery of recombinases [7] and postsynaptic expression of a recombinase-dependent dual gene expression cassette. This system could achieve presynaptic input-defined multi-gene expression in postsynaptic neurons of the S1BF or striatum, where multiple presynaptic innervations are observed.
MATERIALS AND METHODS
Plasmids
pAAV-EF1a-cDIO-EGFP-CMV-fDIO-mScarlet was generated by insertion of the CMV-fDIO-mScarlet fragment from pAAV-CMV-fDIO-mScarlet into the
To produce pAAV-EF1a-cDIO-EGFP, the PCR-amplified coding sequence of EGFP (EGFP-SpeI-S→EGFP-SpeI-A) from pEGFP-C1 (Clontech) was cloned into the
To construct pAAV-EF1a-Cre, the PCR-amplified coding sequence of Cre (Cre-KpnI-S→Cre-EcoRI-A) from pAAV-EF1a-mCherry-IRES-Cre (Addgene #55632) was cloned into the pAAV-EF1a-FlpO plasmid (Addgene #55637). pAAV-hSyn-Cre and pAAV-hSyn-FlpO were obtained by inserting the
The oligonucleotide sequences used in the PCR were as follows: EGFP-SpeI-S, GACTAGTATGGTGAGCAAGGGCGAG; EGFP-SpeI-A, GGACTAGTTTACTTGTACAGCTCGTCC ATG; mScarlet-NheI-S, CTAGCTAGCACCATGGTGAGCAAGGGC; mScarlet-AscI-A, AGGCGCGCC TTACTTGTACAGCTCGTCCATGC; CMV-MluI-S, CGACGCGTTAGTAATCAATTACGGGGTC; CMV-BamHI-A, CGCGGATCCACCGGTAGCGCTAGCG; CMV-EcoRI-S, CCGGAATTCCCGCGT TACATAACTTACGG; WPRE-233n-A, GTTGCGTCAGCAAACACAGT; Cre-KpnI-S, CGGGGTACC ATGGCCAATTTACTGACCG; Cre-EcoRI-A.
Cell culture and transfection
Human embryonic kidney (HEK) 293T cells were cultured in Dulbecco’s modified Eagles medium (DMEM, HyCloneTM) supplemented with 10% fetal bovine serum (FBS, HyCloneTM) at 37°C under 5% CO2. The cultured HEK293T cells were transfected using Lipofectamine 2000 (Invitrogen). After transfection, HEK293T cells were incubated for 48~72 h, depending on the experimental design.
FACS analysis
For fluorescence-activated cell sorting (FACS) analysis, HEK 293T cells were transfected with Cre- and/or FlpO-recombinase and pAAV-EF1a-cDIO-EGFP-CMV-fDIO-mScarlet. After 48 h, cells were harvested with 0.25% trypsin-EDTA (Gibco) and washed with 1X PBS (phosphate buffered saline, pH 7.4), and then centrifuged. The harvested cells were suspended in DMEM medium and analyzed using the Flow Cytometer Cell Sorter (MoFlo Astrios, Beckman Coulter) to evaluate the cross-reactivity of Cre- or FlpO-recombinases. EGFP and mScarlet signals were detected through a 488-513/26 and 561-579/16 nm
Virus injection
Adult mice (C57BL/6J) and
Our dual FP-expression cassette (cGreen-fRed) was packaged in AAV5, which does not show trans-synaptic transfer. AAV5-cGreen-fRed was packaged from the viral vector core at the University of North Carolina (UNC). Stereotaxic coordinates targeting brain areas were as follows (in mm): the primary motor cortex (M1) [anteroposterior (AP)=2.0, mediolateral (ML)=1.8 and dorsoventral (DV; ventral from the cortical surface)=0.5]; the dorsal striatum (dStr) [AP=0.2, ML=2.2, DV=3.4]; the ventral posteromedial nucleus (VPM) [AP=1.8, ML=1.6, and DV=3.6]; the posterior medial nucleus of the thalamus (POm) [AP=2.06, ML=1.25, DV=2.97]; the somatosensory barrel field cortex (S1BF) [AP=0.7, ML=3.5, DV=0.35 and 0.75 (with 30° angle)]; and the vibrissal area of the primary motor cortex (vM1) [AP=1.34, ML=1.5, DV=0.35]. A volume of 200 nl of the virus-containing solution was injected into the M1 and POm with a 6 nl/min injection rate.
Fluorescence imaging and analysis
Fluorescence signals from HEK293T cells were detected using a fluorescent microscope (Nikon eclipse Ts2, objective 40X) equipped with a LED illuminator (excitation wavelength: 470 nm or 525 nm).
To detect fluorescence signals from brain slices, anesthetized mice were dissected and their brains fixed using vascular transcardial perfusion of 1X PBS followed by perfusion of a fixative solution (4% paraformaldehyde in 1X PBS, pH 7.4). Whole brains were then isolated and incubated with the fixative solution for 24 h at 4°C. Fixed brains were then incubated with a 30% sucrose solution (30% sucrose, 1X PBS) for 36~48 h, mounted in frozen section embedding medium (FSC, Leica), and cut at 40 µm thickness using a cryostat (Leica CM 1850 UV Cryostat). Sagittal and coronal slices were prepared for observing and labeling of neurons in the striatum and cortex, respectively. Sections were washed three times in 1X PBS and stained with DAPI (Sigma). After staining, the sections were mounted using a mounting medium (
RESULTS
Validation of AAV1 as a monosynaptic anterograde tracing vector
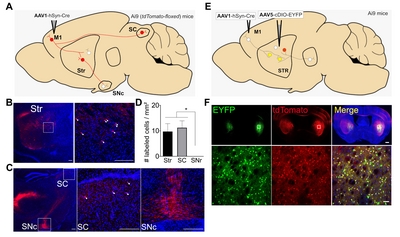
To develop an anterograde trans-synaptic tracing tool for labeling multiple synaptic connections, previously reported anterograde viral vectors such as AAV1 were utilized [7]. AAV1 was selected because of its lower toxicity and easier manipulation than other viral vectors. To confirm if AAV1 was able to perform monosynaptic anterograde transfer from presynaptic to postsynaptic neurons, AAV1-hSyn-Cre was intracranially delivered into the primary motor cortex (M1) of adult
Four weeks after viral injection, neuronal tdTomato expression was found in direct postsynaptic targets of M1, such as the dorsal striatum (dStr) or superior colliculus (SC) [7]. A single bout of AAV1-hSyn-Cre injection (~50 nl of viral particles with a titer of ~1013 genome copy/ml) resulted in similar densities of tdTomato-expressing cells per mm2 in the dStr and SC (number of labeled cells/mm2: dStr=9.7±3.1 vs. SC=11.2±2.7; Fig. 1B~D) in the slices pooled from two mice. However, the substantia nigra pars compacta (SNc), which does not directly receive synaptic inputs from the M1 but is innervated by striatal projecting neurons, only showed limited tdTomato expression in neurite-like structures (Fig. 1C, D). Postsynaptic tdTomato expression in
Designing a dual-gene expression vector for simultaneous anterograde tracing of multiple pathways
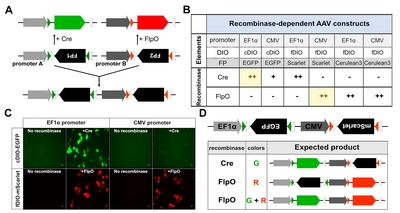
We additionally attempted to develop an anterograde trans-synaptic system that would enable presynaptic input-defined multiple tags in postsynaptic neurons by using AAV1. Postsynaptic expression of multiple tags determined by input connectivity could be achieved by delivering (1) a gene expression cassette of multiple genes (multi-gene expression cassette) into postsynaptic neurons and (2) AAV1, which contains inducers or regulators of gene expression from postsynaptic multi-gene expression cassettes to presynaptic neurons.
To construct a multi-gene expression cassette to be delivered to postsynaptic neurons, we set out to design a dual gene expression cassette for expressing two different fluorescent proteins (FPs). AAV1-mediated delivery of Cre recombinase from presynaptic to postsynaptic neurons to induce expression of FP genes flanked by DIO is possible (Fig. 2A). Moreover, regulated expression of two different FP genes could be achieved if each FP gene was flanked with different DIO elements, and corresponding recombinases were delivered from presynaptic inputs via AAV1. By searching for recombinases that are selective and efficient for the expression of DIO-flanked FPs and promoters ideal for FP expression, we constructed a series of mammalian expression vectors containing strong general promoters such as human elongation factor 1a promoter (EF1a), cytomegalovirus immediate-early promoter (CMV) or human ubiquitin C promoter (UbC), combined with a recombinase-dependent gene expression cassette (Fig. 2B). Previous studies have shown that site-specific recombinases such as Cre, FlpO, and vCre recognize specific DNA sequences such as loxP/lox2272, FRT/F5, and vloxP/vlox2272, respectively [9, 10]. Inverse-oriented gene sequences encoding FPs with different emission wavelengths such as EGFP (Green), mScarlet (Red) or mCerulean3 (Blue) were flanked with recombinase-specific DIO sequences, resulting in cDIO-, fDIO (FlpO-responsive DIO)-, and vDIO (vCre-responsive DIO)- FPs (Fig. 2B).
After transfecting plasmids containing different combinations of promoters, reporters, and DIOs into HEK293T cells, we then tested which construct was most efficient and selective for FP expression (Fig. 2C). Consistent with previous reports, cDIO- or fDIO-FP expression did not overlap when each recombinase was combined (Fig. 2B, C). In addition, our data showed that the EF1α promoter is more efficient in expressing cDIO-EGFP than the CMV promoter (Fig. 2B, C). However, the CMV promoter with fDIO was suitable for high expression of mScarlet when floxed in fDIO (Fig. 2B, C). Since both EGFP and mScarlet showed brighter expression than the blue fluorescent protein mCerulean3, we selected EF1α-cDIO-EGFP and CMV-fDIO-mScarlet cassettes for the dual FP expression cassette. A fusion construct including both cassettes was inserted into the single AAV genome, resulting in AAV-EF1α-cDIO-EGFP-CMV-fDIO-mScarlet (AAV-cGreen-fRed; Fig. 2D).
Cre and/or FlpO-dependent EGFP and mScarlet expression from a dual-gene expression vector
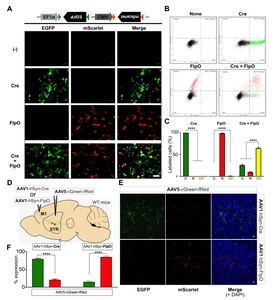
To test whether the newly developed dual gene expression cassette in the AAV vector was able to express EGFP and/or mScarlet by selective actions of Cre or FlpO recombinases, AAV5-cGreen-fRed was introduced into HEK293T cells with Cre and/or FlpO expression (Fig. 3A). Our data showed that expression of either EGFP or mScarlet was exclusively observed in the presence of the corresponding recombinase, and co-expression of Cre and FlpO was sufficient for dual expression of EGFP and mScarlet (Fig. 3A). When EGFP or mScarlet expression was quantitatively assessed by flow cytometry analysis of transfected HEK293T cells, we found that 99.96±0.02% of cells showed EGFP expression when Cre was present, and 99.02±0.25% of FlpO-expressing cells were sorted as mScarlet expressing cells (from three different batches of cell cultures; Fig. 3B, C). Nearly 65% of cells displayed co-expression of EGFP and mScarlet by co-transfection of Cre and FlpO (EGFP only: 25.68±1.37%; mScarlet only: 9.65±0.97%, EGFP+mScarlet: 64.67±2.20%; from three different batches of cell cultures; Fig. 3B, C). These results indicate that our dual gene expression cassette efficiently expressed multiple FPs, and that the expression of these FPs was selectively regulated by the corresponding Cre or FlpO recombinases.
To verify recombinase-specific FP expression from AAV5-cGreen-fRed
In vivo dual color labeling of postsynaptic neurons receiving multiple inputs
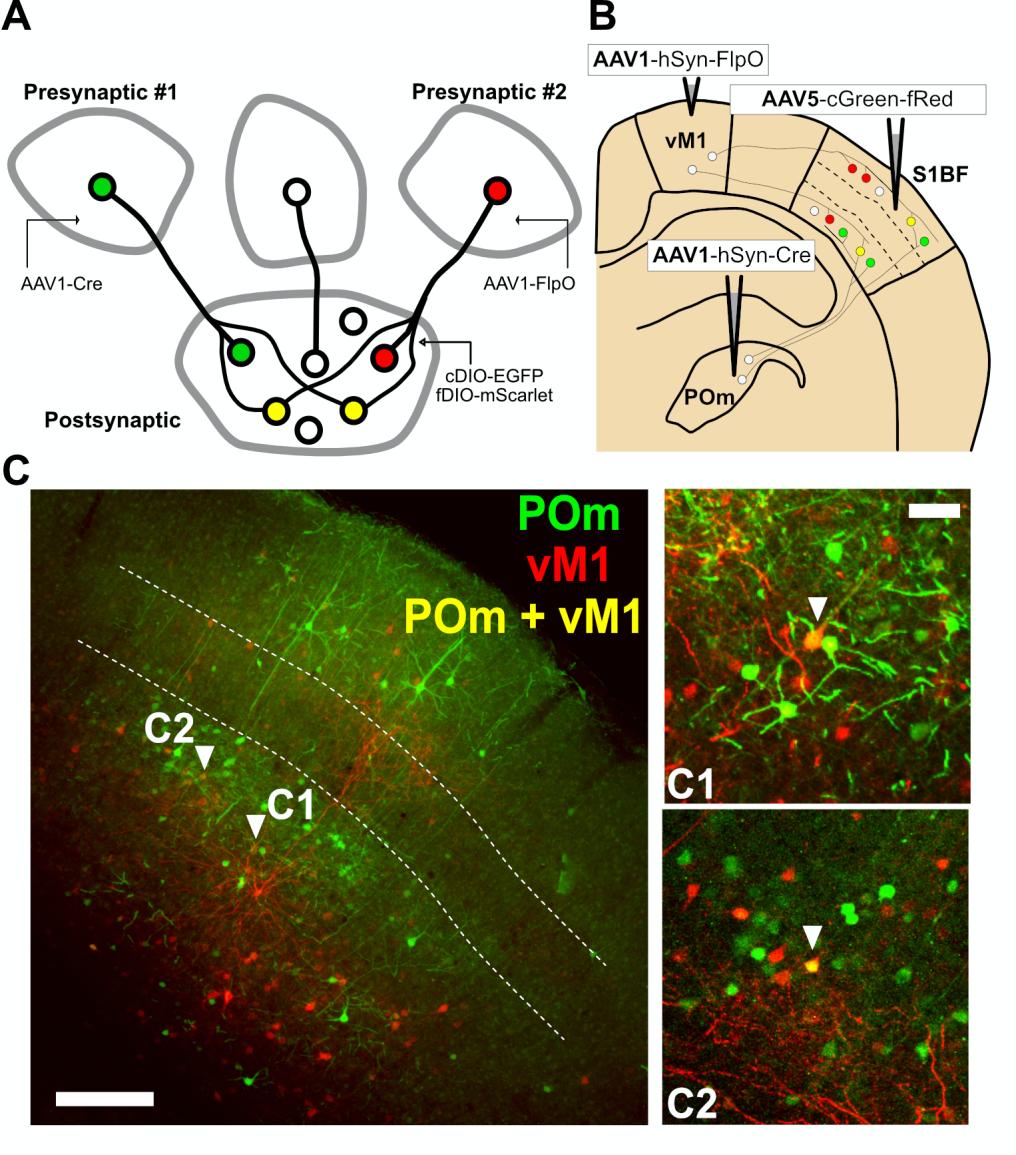
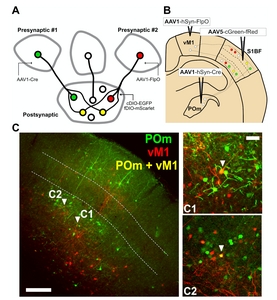
Since EGFP or mScarlet expression can be individually controlled by Cre or FlpO in the same cell when our dual gene expression cassette was introduced, we next tested whether AAV1-mediated trans-synaptic transfer of Cre or FlpO from presynaptic neurons can regulate EGFP or mScarlet expression in postsynaptic neurons with AAV-cGreen-fRed (Fig. 4A).
We first took advantage of the canonical wiring selectivity of the somatosensory barrel field cortex (S1BF). The posterior medial nucleus of the thalamus (POm) is known to primarily innervate layer 1 (L1) and 5a (L5a), but avoids layer 4 (L4) of the S1BF [11, 12]. Likewise, S1BF receives presynaptic inputs from higher-order cortical inputs such as presynaptic innervation from the vibrissal area of the primary motor cortex (vM1), which also projects mainly on infragranular layers, yet scarcely on L4 [11, 13, 14]. We thus tested whether S1BF neurons could be defined by cells receiving presynaptic inputs from POm (EGFP-positive) or vM1 (mScarlet-positive). AAV1-hSyn-Cre and -FlpO were injected into presynaptic POm and vM1, respectively, and AAV5- cGreen-fRed into the postsynaptic S1BF (Fig. 4B). Four weeks after injection, we found that POm or M1-receiving neurons were mainly observed in L2/3, L5, and L6, but scarcely in L4 in the S1BF (Fig. 4C)—corroborating previous findings [11,15,-17].
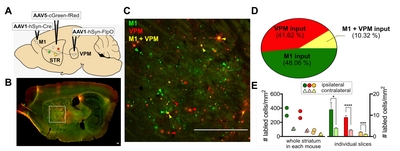
Next, our cFork system was tested in striatal circuits of the mouse brain (Fig. 5A). Because the dorsal striatum receives two major excitatory inputs from the cortex and thalamus, called the corticostriatal and thalamostriatal pathways, respectively [18,-20], we aimed to label striatal postsynaptic neurons with EGFP and/or mScarlet, whose expression is defined by presynaptic cortical and/or thalamic inputs, respectively. For this purpose, AAV1s containing Cre or FlpO were injected into the right hemisphere of the M1 cortical subregion or into a part of the ventral posteromedial (VPM) thalamic nucleus of wild-type C57/B6 mice. Bilateral dStr was injected with AAV5-cGreen-fRed (Fig. 5A).
Four weeks after viral injection, we found mixed patterns of EGFP and/or mScarlet expression in striatal neurons (Fig. 4B, C). These FP expression patterns seemed to be mediated by anterograde transport of Cre and FlpO from M1 and VPM, respectively, because no FP expression was observed in the striatum when AAV1-hSyn-Cre or hSyn-FlpO was not injected (data not shown). The percentage of cells showing exclusive expression of EGFP and mScarlet was 48.06% (4,233/8,808 cells) and 41.62% (3,666/8,808 cells), respectively, and 10.32% (909/8,808 cells) of striatal cells showed co-expression of EGFP and mScarlet (whole striatal areas derived from sagittal sections from two mice; Fig. 5D). Despite estimation from two animals and an 80% accuracy of FP-labeling by presynaptic input-derived recombinases (Fig. 3F), our results (Fig. 5D) are in line with previous findings that striatal medium spiny neurons contain corticostriatal and thalamostriatal synapses equally [21, 22], and that projections from M1 and VPM show low convergence in the striatum [20]. Moreover, we found that FP-labeled ipsilateral striatal neurons were approximately four fold higher in number than contralateral ones (ipsilateral vs. contralateral projection shown as number of labeled cells/mm2: M1-input=12.7±3.3 vs. 3.9±0.3, VPM-input=9.0±0.8 vs. 3.0±0.3; n=57 ~ 74 slices from 2 mice; Fig. 5E). These results indicate that ipsilateral corticostriatal and thalamostriatal projections are dominant in the dStr, as previously reported [22,-24].
Together, these results support the idea that cFork is useful for achieving labeling of postsynaptic neurons with two different input-defined tags in the mouse brain.
DISCUSSION
Tracing multiple synaptic connections is essential for understanding how the neural connectome and its activity regulate brain functions. Available neuroanatomical tracing tools can efficiently dissect and identify input/output relationships of two brain regions but cannot effectively address complex and multiple connectivity. Our newly developed anterograde viral tracing system is capable of expressing two different labeling genes in postsynaptic neurons. Expression of these genes was tightly regulated by different recombinases
Despite the presented advantages, there are several points for improvement when designing anterograde viral vectors ideal for multi-connectivity detection in future studies. First, as shown in Fig. 3D-F, recombinase-specific regulation of gene expression from our dual gene expression cassette was ideal
To overcome these limitations, several approaches may be possible. Presynaptic input-dependent tight regulation of gene expression from the dual-gene expression vector may be achieved by re-designing and screening of more specific DIO and FP combinations, which selectively respond to the AAV1-mediated anterograde delivery of recombinases. Experimental artifacts and the small genome capacity of AAV could be overcome if transgenic/knock-in mice constitutively expressing multi-gene expression cassettes are constructed. Moreover, along with efforts to identify additional specific recombinases-DIO pairs, novel anterograde viral vectors which are easy to handle and have a larger genome capacity than AAV may facilitate the development of an advanced version of the anterograde tracing tool, thus enabling simultaneous visualization of multi-synaptic connectivity in the postsynaptic neuron.
In summary, we have provided a new anterograde tracing tool that can simultaneously visualize dual presynaptic input connectivity in postsynaptic neurons based on the anterograde AAV vector. Our tracing system is an ideal viral tracing tool capable of dissecting multiple synaptic connectivity with minimal effort.
ACKNOWLEDGEMENTS
This work was supported by the KBRI basic research program (19-BR-04-01, 19-BR-01-03) of the Korea Brain Research Institute, and Brain Research Program (NRF-2017M3C7A1048086) of the National Research Foundation of Korea (NRF), funded by the Ministry of Science and ICT.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
- Lanciego JL, Wouterlood FG (2011) A half century of experimental neuroanatomical tracing. J Chem Neuroanat 42: 157-183


- Nassi JJ, Cepko CL, Born RT, Beier KT (2015) Neuroanatomy goes viral!. Front Neuroanat 9: 80

- Ugolini G, Kuypers HG, Strick PL (1989) Transneuronal transfer of herpes virus from peripheral nerves to cortex and brainstem. Science 243: 89-91
- Callaway EM (2008) Transneuronal circuit tracing with neurotropic viruses. Curr Opin Neurobiol 18: 617-623
- Lo L, Anderson DJ (2011) A Cre-dependent, anterograde transsynaptic viral tracer for mapping output pathways of genetically marked neurons. Neuron 72: 938-950
- Beier KT, Saunders A, Oldenburg IA, Miyamichi K, Akhtar N, Luo L, Whelan SPJ, Sabatini B, Cepko CL (2011) Anterograde or retrograde transsynaptic labeling of CNS neurons with vesicular stomatitis virus vectors. Proc Natl Acad Sci U S A 108: 15414-15419
- Zingg B, Chou XL, Zhang ZG, Mesik L, Liang F, Tao HW, Zhang LI (2017) AAV-mediated anterograde transsynaptic tagging: mapping corticocollicular input-defined neural pathways for defense behaviors. Neuron 93: 33-47
- Jhang J, Lee H, Kang MS, Lee HS, Park H, Han JH (2018) Anterior cingulate cortex and its input to the basolateral amygdala control innate fear response. Nat Commun 9: 2744
- Minorikawa S, Nakayama M (2011) Recombinase-mediated cassette exchange (RMCE) and BAC engineering via VCre/VloxP and SCre/SloxP systems. Biotechniques 50: 235-246
- Fenno LE, Mattis J, Ramakrishnan C, Hyun M, Lee SY, He M, Tucciarone J, Selimbeyoglu A, Berndt A, Grosenick L, Zalocusky KA, Bernstein H, Swanson H, Perry C, Diester I, Boyce FM, Bass CE, Neve R, Huang ZJ, Deisseroth K (2014) Targeting cells with single vectors using multiple-feature Boolean logic. Nat Methods 11: 763-772
- Petreanu L, Mao T, Sternson SM, Svoboda K (2009) The subcellular organization of neocortical excitatory connections. Nature 457: 1142-1145
- Aronoff R, Matyas F, Mateo C, Ciron C, Schneider B, Petersen CC (2010) Long-range connectivity of mouse primary somatosensory barrel cortex. Eur J Neurosci 31: 2221-2233
- Veinante P, Deschênes M (2003) Single-cell study of motor cortex projections to the barrel field in rats. J Comp Neurol 464: 98-103
- Petersen CC (2007) The functional organization of the barrel cortex. Neuron 56: 339-355
- Jouhanneau JS, Ferrarese L, Estebanez L, Audette NJ, Brecht M, Barth AL, Poulet JF (2014) Cortical fosGFP expression reveals broad receptive field excitatory neurons targeted by POm. Neuron 84: 1065-1078
- Kinnischtzke AK, Simons DJ, Fanselow EE (2014) Motor cortex broadly engages excitatory and inhibitory neurons in somatosensory barrel cortex. Cereb Cortex 24: 2237-2248
- Zhang W, Bruno RM (2019) High-order thalamic inputs to primary somatosensory cortex are stronger and longer lasting than cortical inputs. Elife 8: e44158
- Pan WX, Mao T, Dudman JT (2010) Inputs to the dorsal striatum of the mouse reflect the parallel circuit architecture of the forebrain. Front Neuroanat 4: 147
- Hunnicutt BJ, Long BR, Kusefoglu D, Gertz KJ, Zhong H, Mao T (2014) A comprehensive thalamocortical projection map at the mesoscopic level. Nat Neurosci 17: 1276-1285
- Hunnicutt BJ, Jongbloets BC, Birdsong WT, Gertz KJ, Zhong H, Mao T (2016) A comprehensive excitatory input map of the striatum reveals novel functional organization. Elife 5: e19103
- Ding J, Peterson JD, Surmeier DJ (2008) Corticostriatal and thalamostriatal synapses have distinctive properties. J Neurosci 28: 6483-6492
- Smith Y, Raju DV, Pare JF, Sidibe M (2004) The thalamostriatal system: a highly specific network of the basal ganglia circuitry. Trends Neurosci 27: 520-527
- Carman JB, Cowan WM, Powell TP, Webster KE (1965) A bilateral cortico-striate projection. J Neurol Neurosurg Psychiatry 28: 71-77
- Donoghue JP, Herkenham M (1986) Neostriatal projections from individual cortical fields conform to histochemically distinct striatal compartments in the rat. Brain Res 365: 397-403
- Warnock JN, Daigre C, Al-Rubeai M (2011) Introduction to viral vectors. Methods Mol Biol 737: 1-25