Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2019; 28(5): 602-611
Published online October 31, 2019
https://doi.org/10.5607/en.2019.28.5.602
© The Korean Society for Brain and Neural Sciences
Combination of In Vivo [123I]FP-CIT SPECT and Microdialysis Reveals an Antipsychotic Drug Haloperidol-induced Synaptic Dopamine Availability in the Rat Midbrain and Striatum
So Hyeon Park1, Yoo Sung Song2, Byung Seok Moon3, Byung Chul Lee2, Hyun Soo Park1,2* and Sang Eun Kim1,2,4*
1Department of Transdisciplinary Studies, Graduate School of Convergence Science and Technology, Seoul National University, Seoul 08826, 2Department of Nuclear Medicine, Seoul National University Bundang Hospital, Seoul National University College of Medicine, Seoul 03080, 3Department of Nuclear Medicine, Ewha Woman’s University Seoul Hospital, Ewha Womans University College of Medicine, Seoul 07804, 4Advanced Institutes of Convergence Technology, Suwon 16229, Korea
Correspondence to: *To whom correspondence should be addressed.
Sang Eun Kim, TEL: 82-31-787-7671, FAX: 82-31-787-4018
e-mail: kse@snu.ac.kr
Hyun Soo Park, TEL: 82-31-787-2936, FAX: 82-31-787-4018
e-mail: hyuns@snu.ac.kr
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License
(http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and
reproduction in any medium, provided the original work is properly cited.
Abstract
Synaptic dopamine (DA) is mainly regulated by the presynaptic DA transporter (DAT). Single-photon emission computerized tomography (SPECT) with the DAT radiotracer [123I]FP-CIT assesses changes in synaptic DA availability when endogenous DA displaces [123I]FP-CIT or competes for DAT. Here, we investigated the effects of haloperidol (HAL) and clozapine (CLZ) on [123I]FP-CIT binding in the rat striatum and midbrain to assess the utility of [123I]FP-CIT SPECT to quantify changes in synaptic DA availability. Rats underwent [123I]FP-CIT SPECT after intraperitoneal administration of normal saline (vehicle), HAL (1 and 7 mg/kg), CLZ (10 and 54 mg/kg) and bupropion (BUP, a DAT blocker, 20 and 100 mg/kg). In the striatum and midbrain, percent differences in the nondisplaceable binding potential (BPND) of [123I]FP-CIT compared to the vehicle were calculated for the various drugs and doses. In another experiment, changes in endogenous striatal DA concentration were measured by
Graphical Abstract
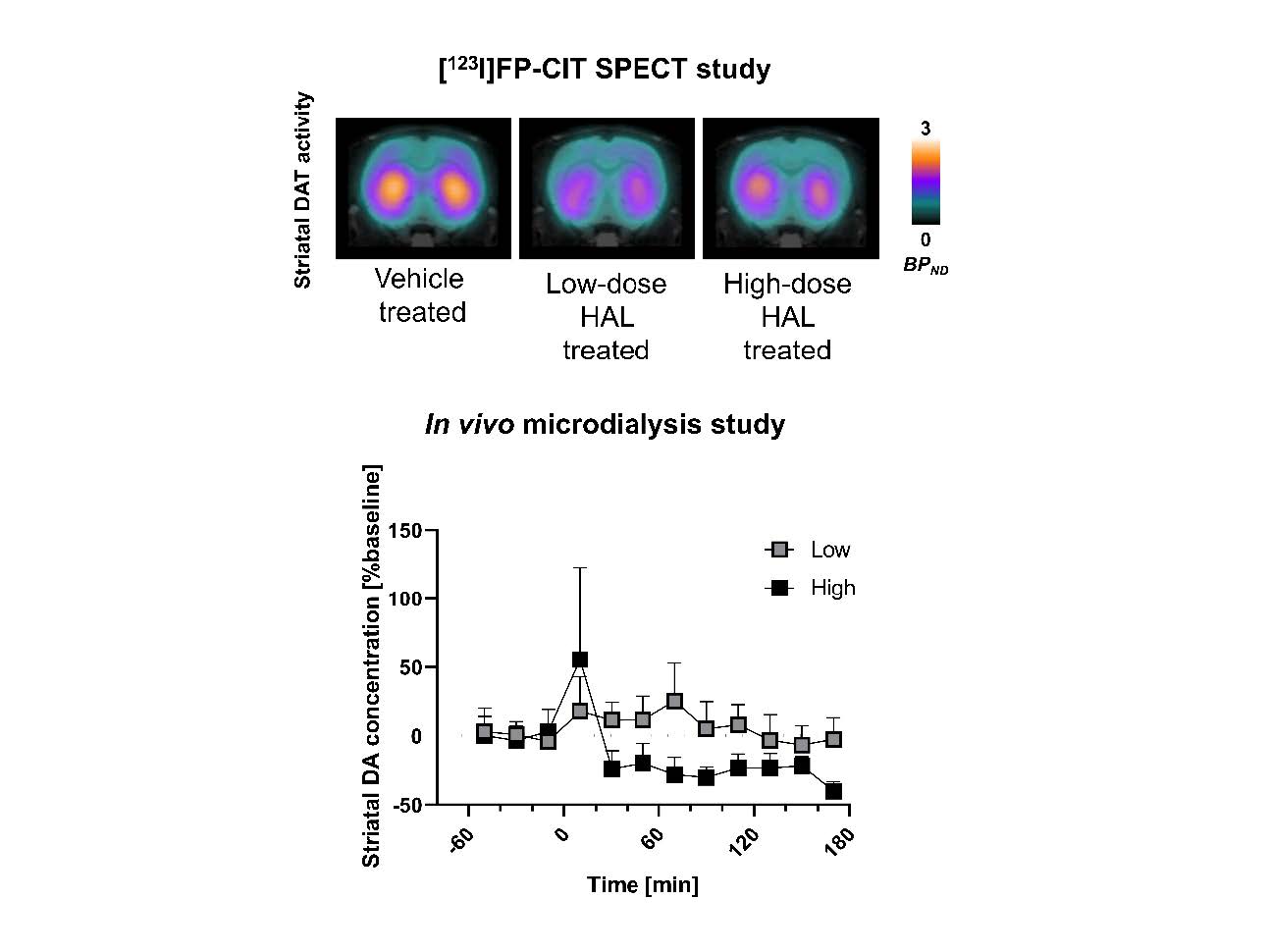
Keywords: [123I]FP-CIT SPECT, Dopamine availability, Haloperidol, Clozapine,
INTRODUCTION
Synaptic dopamine (DA) availability is implicated in the pathology of various neurological and psychiatric diseases, such as Parkinson’s disease [1, 2] and schizophrenia [3, 4]. DA availability in the brain is mainly regulated by the DA transporter (DAT) [5], and DA receptors for its D2 and D3 subtypes expressed on the cell membrane of DA neurons (i.e., autoreceptors) also play a key role in regulating the activity of DAergic neurons and controlling DA synthesis, release, and reuptake [6]. Under the DA hypothesis, antipsychotic drugs act mainly by regulating the DAergic systems, as evidenced by the significant association between DA receptor antagonism and improved positive and/or negative symptoms. Some of them have been proven to regulate DA release [6–8]. An antipsychotic drug haloperidol (HAL) is known to increase DA synthesis and release in the striatum and related mesolimbic structures [9–12]. The proposed mechanism of action consists of presynaptic terminal blockade by DA autoreceptors [11], which abolishes feedback inhibition, leading to enhanced DA synthesis or release. Clozapine (CLZ), despite acting as a DA antagonist as part of its therapeutic effect against schizophrenia symptoms, also stimulates DA release in the ventral striatum (i.e., nucleus accumbens) of rats [13] and in the hippocampus of schizophrenia patients [14]. Presynaptic modulation induced by antipsychotic drugs affects the extracellular DA concentration by altering autoregulation [15]. An increased extracellular DA concentration can lead to stimulation of DA autoreceptors, which inhibits DA release [6, 16]. DA release and metabolism in the rat striatum
The competition between endogenous transmitters and radio-labeled ligands for
We investigated differences in the nondisplaceable binding potential of [123I]FP-CIT in the rat striatum and midbrain, which compose the nigrostriatal DA system, after HAL and CLZ compared to vehicle treatment in an attempt to assess the validity of [123I]FP-CIT SPECT as a measure of synaptic DA availability induced by HAL and CLZ. The reliability of this noninvasive imaging technique was further examined for the striatum by conducting an
MATERIALS AND METHODS
Animals and drugs
This study was approved by the Institutional Animal Care and Use Committee of the Seoul National University Bundang Hospital. Animals were purchased from Orient Bio Inc., Seoul, Korea. A total of 35 and 20 Sprague Dawley (SD) rats (male, 6-week-old, 260~300 g body weight) were used for [123I]FP-CIT SPECT and
HAL, CLZ and bupropion (BUP) hydrochloride were purchased from Sigma-Aldrich Korea, Yongin, Korea. HAL and CLZ were dissolved in 1% tartaric acid and 1 N HCl. A DAT blocker, BUP, the positive control drug for HAL and CLZ, was used to test whether endogenous DA displaces [123I]FP-CIT or BUP competes with [123I] FP-CIT for DAT. BUP was dissolved in NS.
In the [123I]FP-CIT SPECT study, the animals were divided into four groups defined by different drugs (vehicle (n=5), HAL (n=10), CLZ (n=10), BUP (n=10) treatment groups). While the vehicle-treated group had no dose conditions, the others had low and high dose conditions for the drug treatment. The low and high doses of HAL, CLZ and BUP were 1 and 7 mg/kg body weight, 10 and 54 mg/kg body weight, and 20 and 100 mg/kg body weight, respectively (n=5 per drug and dose condition). The doses were selected based on previous studies showing that low and high doses induce ~25% increases in synaptic DA availability and greater than 80% DA receptor occupancy by drugs in the striatum, respectively [24–27].
In another experiment, changes in endogenous DA concentration in the striatum were monitored by
In both experiments, drugs were injected intraperitoneally (
[123I]FP-CIT SPECT/CT study
The [123I]FP-CIT SPECT/CT study was performed on a dedicated small-animal SPECT/CT system (NanoSPECT/CT, Mediso Inc., Budapest, Hungary). Helical small-animal SPECT scans were performed using a 4-head γ-camera outfitted with multipinhole collimators (1.4-mm-diameter pinholes) designed for rats. [123I] FP-CIT was injected at a dose (mean±SD) of 39.5±7.2 MBq 1 h after drug treatment; 2 h later (once [123I]FP-CIT had reached equilibrium in the striatum), SPECT/CT data were acquired from the animals for 30 min under 2% isoflurane anesthesia. After the scan, the SPECT data were reconstructed using iterative three-dimensional ordered subset expectation maximization with the single-slice rebinning method. CT-based attenuation correction was performed, as were scatter and random correction. The reconstructed images were 176×176×136 pixels with a voxel size of 0.6×0.6×0.6 mm (x, y, z). PMOD software (PMOD Technologies LLC., Geneva, Switzerland) was used for processing and analysis of SPECT and CT images. Images were spatially normalized to standard stereotaxic space with the predefined magnetic resonance imaging (MRI) rat brain template. The striatum, midbrain and cerebellum were defined using automated anatomical labeling embedded in PMOD software [28]. Synaptic DA availability in the striatum and midbrain was quantitatively assessed in terms of the nondisplaceable binding potential (BPND) of [123I]FP-CIT, which is proportional to the density of available binding sites (i.e., DAT). The cerebellum (which is known as a DAT-poor region or nondisplaceable binding site of [123I]FP-CIT) was set as the reference region for estimating BPND according to the following equation: BPND=(
In vivo microdialysis study
Extracellular DA concentrations in the striatum of freely moving rats were directly measured by
Statistical analysis
Data were analyzed using GraphPad Prism (version 7.0, Graph-Pad Software Inc., La Jolla, CA, USA). The statistical significance of differences in mean BPND between drug and dose conditions was tested by two-way analysis of variance (ANOVA), followed by Tukey’s multiple comparison test between dose conditions for each drug.
RESULTS
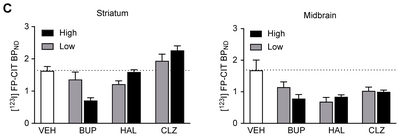
In the vehicle-treated group, the mean [123I]FP-CIT BPND was 1.64±0.13 and 1.69±0.32 in the striatum and midbrain, respectively; these values are comparable to the results of previous studies [24, 30]. BUP dose-dependently occupied DAT to a considerable degree, as evidenced by decreases in [123I]FP-CIT BPND of −16.50% (20 mg/kg) and −56.29% (100 mg/kg) in the striatum and −31.57% (20 mg/kg) and −53.08% (100 mg/kg) in the midbrain, implying that [123I]FP-CIT SPECT is a reliable and sensitive technique for measuring drug-induced changes in DAT activity and can conceptually allow the assessment of changes in synaptic DA availability
Treatment with HAL and CLZ markedly altered synaptic DA availability compared to the vehicle, as evidenced by changes in [123I]FP-CIT BPND in both the striatum and the midbrain. Intriguingly, the level of changes in [123I]FP-CIT BPND treatment varied across drugs, doses, and regions (Fig. 1 and Table 1). Compared to the vehicle, HAL decreased [123I]FP-CIT BPND in the striatum (−25.29% and −2.27% for 1 and 7 mg/kg, respectively) and to a greater degree in the midbrain (−58.74% and −49.64% for 1 and 7 mg/kg, respectively), whereas the CLZ-treated group showed an increase in the striatum (18.85% and 38.64% for 10 and 54 mg/kg, respectively) but a decrease in the midbrain (−38.60% and −40.38% for 10 and 54 mg/kg, respectively).
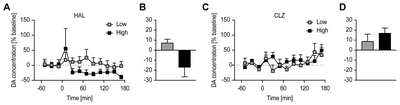
The changes in extracellular striatal DA concentrations by HAL (1 and 7 mg/kg) and CLZ (10 and 54 mg/kg) treatment were evaluated by
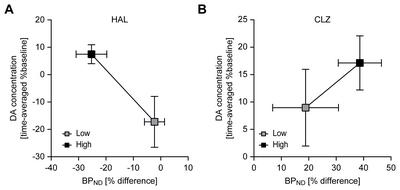
The relationships between percentage differences in [123I]FP-CIT BPND and time-averaged percentage changes from baseline extracellular DA concentration after treatment with varying doses of HAL and CLZ are depicted in Fig. 3 (A and B for HAL and CLZ, respectively). An inverse relationship between percentage differences in [123I]FP-CIT BPND and time-averaged % percentage change from baseline extracellular DA concentration (lower [123I] FP-CIT BPND, greater synaptic DA availability) appeared only in the HAL-treated group.
DISCUSSION
Under the investigational assumption that if endogenous DA displaces radioligands or competes with them for presynaptic binding sites, radioligand binding to the DAT could be affected, we investigated differences in the [123I]FP-CIT BPND in the rat striatum and midbrain, which compose the nigrostriatal DA system, after HAL and CLZ treatments that increased synaptic DA availability. The reliability of this noninvasive imaging technique was further examined for the striatum by using an
Our results showed that [123I]FP-CIT SPECT allows the detection of apparent increases in synaptic DA concentration induced by low (1 mg/kg
The [123I]FP-CIT SPECT and
Although it is possible that there were experimental errors and unknown contamination factors that could have led to these results, the magnitude of error in the measured values (BPND and DA concentration) was quite acceptable. The reliability of increased striatal [123I]FP-CIT BPND could be validated from decreases in the midbrain of the same subject on CLZ treatment. In the midbrain, non-dose-dependent decreases in [123I]FP-CIT binding after treatment were consistently shown among drugs (although the DAT blocker BUP occupied DAT dose-dependently). Regardless of dose, the %difference was greater for HAL (−49.64~58.74% difference) than CLZ (−38.60 to −40.38% difference) in accordingly with pharmacologic characteristics in DA regulation of HAL and CLZ, implying their own typicality. That CLZ affects increasing binding affinity to DAT is unlikely, but it is not impossible, as some ligands occasionally act in this way. For example, an antiepileptic drug, tiagabine, which binds to the central benzodiazepine receptor, affects the increased binding affinity of radiolabeled ligand ([18F] flumazenil) to the central benzodiazepine receptor, known as the “GABA shift” [35]. On the other hand, the interaction between CLZ and [123I]FP-CIT could also be considered. The potential effect of CLZ on the group of antipsychotic drugs was examined; for instance, only CLZ induced a decrease in the protein kinase C level [36], but the effect of decreased protein kinase C level and interaction between [123I]FP-CIT and DAT is unexpected.
In addition, this study examined changes in synaptic DA availability after acute administration of HAL and CLZ in an attempt to resolve the previously reported controversy [12, 13] through an analogous approach to the experimental paradigm and to determine whether [123I]FP-CIT SPECT can be used to assess changes in endogenous DA concentration based on alterations in [123I]FP-CIT BPND to DAT as it is displaced by endogenous DA. Importantly, we partly overcame the experimental limitations of the earlier studies by performing
DAT imaging is increasingly used for diagnostics and drug development for neurological and psychiatric diseases as well as for precision medicine in cases of complicated medication status—for instance, a Parkinson’s disease patient receiving antipsychotic drugs. Consequently, there is a need for further experimental evidence supporting the utility of [123I]FP-CIT SPECT for quantitation of acute changes in available DA. Recently, there has been growing interest in assessing DAT binding in schizophrenia patients. However, the results obtained to date on striatal DAT binding in schizophrenia subjects have been inconsistent, with reports of elevated [37, 38], reduced [22, 38, 39], or unaltered [23, 40–44] DAT binding. Interestingly, unaltered [43], decreased [22, 39], or increased [37] DAT binding has also been observed in medicated patients. These results are difficult to interpret because factors such as illness duration and phase–which could vary between patients and investigations—are likely to affect the regulation of pre- and postsynaptic binding sites. Moreover, our findings suggest that antipsychotic drugs themselves may confound presynaptic binding data. Schizophrenia patients who are not responding to antipsychotic drug treatment can have a high percentage of occupied D2 receptors without any relief of symptoms [45]. In light of the present findings, it is conceivable that presynaptic autoreceptor or transporter function may be dysregulated in this subgroup of schizophrenia patients. In routine clinical studies as well as scientific studies, patients are frequently on medication and sometimes even take drugs of abuse [46]. Moreover, in preclinical studies, animals are anesthetized for their scans. Prescribed drugs, drugs of abuse, and anesthetics may influence the visual interpretation and/or quantification of [123I]FP-CIT SPECT scans.
The present study has several limitations. We used different methods to measure alterations in the DAergic neurotransmission system induced by treatment with the antipsychotic drugs HAL and CLZ. The two techniques employed have pros and cons with respect to their ability to measure alterations in the DAergic neurotransmission system, and they were used under fundamentally different experimental conditions (anesthetized animals vs. awake animals). Nonetheless, we felt that the utility of [123I]FP-CIT SPECT compared to the
In conclusion, this study demonstrates that [123I]FP-CIT SPECT may be a useful preclinical technique for detecting increases in synaptic DA availability induced by HAL treatment in both the midbrain and the striatum, with results comparable to those obtained by
ACKNOWLEDGEMENTS
This study was supported by the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (HI16C-0947), and by the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT, and Future Planning, Republic of Korea (NRF-2018R1D1A1B07047994, 2016R1D1A1A02937028).
Figures
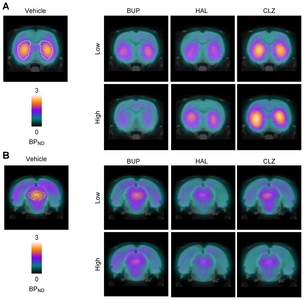
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Tables
Table 1 Differences in [123I]FP-CIT BPND
| Drug | Dose (mg/kg) | Striatum | Midbrain | ||||
|---|---|---|---|---|---|---|---|
| BPND | % Difference | p | BPND | % Difference | p | ||
| VEH | - | 1.64±0.13 | - | - | 1.69±0.32 | - | - |
| BUP | 20 | 1.37±0.22 | −16.50±13.41 | 0.3954 | 1.16±0.16 | −31.57±9.26 | 0.2165 |
| 100 | 0.72±0.08 | −56.29±4.59 | 0.0006 | 0.79±0.12 | −53.08±7.21 | 0.0376 | |
| HAL | 1 | 1.22±0.09 | −25.29±5.60 | 0.0749 | 0.70±0.12 | −58.74±7.38 | 0.0041 |
| 7 | 1.60±0.06 | −2.27±3.65 | 0.9797 | 0.85±0.05 | −49.64±3.13 | 0.0215 | |
| CLZ | 10 | 1.95±0.20 | 18.85±12.00 | 0.2257 | 1.04±0.11 | −38.60±6.53 | 0.0668 |
| 54 | 2.27±0.13 | 38.64±7.87 | 0.0061 | 1.01±0.05 | −40.38±2.76 | 0.0676 | |
Values are the mean±SEM. VEH, vehicle; BUP, bupropion; HAL, haloperidol; CLZ, clozapine.
References
- Damier P, Hirsch EC, Agid Y, Graybiel AM (1999) The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson's disease. Brain 122(Pt 8):1437-1448.


- Joe EH, Choi DJ, An J, Eun JH, Jou I, Park S (2018) Astrocytes, microglia, and Parkinson's disease. Exp Neurobiol 27:77-87.

- Stevens JR (1979) Schizophrenia and dopamine regulation in the mesolimbic system. Trends Neurosci 2:102-105.
- Seeman P, Kapur S (2000) Schizophrenia: more dopamine, more D2 receptors. Proc Natl Acad Sci U S A 97:7673-7675.
- Vaughan RA, Foster JD (2013) Mechanisms of dopamine transporter regulation in normal and disease states. Trends Pharmacol Sci 34:489-496.
- Schmitz Y, Benoit-Marand M, Gonon F, Sulzer D (2003) Presynaptic regulation of dopaminergic neurotransmission. J Neurochem 87:273-289.
- Ichikawa J, Meltzer HY (1991) Differential effects of repeated treatment with haloperidol and clozapine on dopamine release and metabolism in the striatum and the nucleus accumbens. J Pharmacol Exp Ther 256:348-357.
- Starke K, Späth L, Wichmann T (1984) Effects of verapamil, diltiazem and ryosidine on the release of dopamine and acetylcholine in rabbit caudate nucleus slices. Naunyn Schmiedebergs Arch Pharmacol 325:124-130.
- Wu Q, Reith ME, Walker QD, Kuhn CM, Carroll FI, Garris PA (2002) Concurrent autoreceptor-mediated control of dopamine release and uptake during neurotransmission: an in vivo voltammetric study. J Neurosci 22:6272-6281.
- Pehek EA, Yamamoto BK (1994) Differential effects of locally administered clozapine and haloperidol on dopamine efflux in the rat prefrontal cortex and caudate-putamen. J Neurochem 63:2118-2124.
- Pehek EA (1999) Comparison of effects of haloperidol administration on amphetamine-stimulated dopamine release in the rat medial prefrontal cortex and dorsal striatum. J Pharmacol Exp Ther 289:14-23.
- Moghaddam B, Bunney BS (1990) Acute effects of typical and atypical antipsychotic drugs on the release of dopamine from prefrontal cortex, nucleus accumbens, and striatum of the rat: an in vivo microdialysis study. J Neurochem 54:1755-1760.
- Shilliam CS, Dawson LA (2005) The effect of clozapine on extracellular dopamine levels in the shell subregion of the rat nucleus accumbens is reversed following chronic administration: comparison with a selective 5-HT(2C) receptor antagonist. Neuropsychopharmacology 30:372-380.
- Chung YC, Park IS, Li Z, Dai J, Meltzer HY, Ichikawa J (2003) Clozapine, but not haloperidol, increases hippocampal dopamine and acetylcholine release. Clin Psychopharmacol Neurosci 1:79-85.
- Missale C, Nash SR, Robinson SW, Jaber M, Caron MG (1998) Dopamine receptors: from structure to function. Physiol Rev 78:189-225.
- Benoit-Marand M, Borrelli E, Gonon F (2001) Inhibition of dopamine release via presynaptic D2 receptors: time course and functional characteristics in vivo. J Neurosci 21:9134- 9141.
- Gainetdinov RR, Grekhova TV, Sotnikova TD, Rayevsky KS (1994) Dopamine D2 and D3 receptor preferring antagonists differentially affect striatal dopamine release and metabolism in conscious rats. Eur J Pharmacol 261:327-331.
- Westerink BH, Kawahara Y, De Boer P, Geels C, De Vries JB, Wikström HV, Van Kalkeren A, Van Vliet B, Kruse CG, Long SK (2001) Antipsychotic drugs classified by their effects on the release of dopamine and noradrenaline in the prefrontal cortex and striatum. Eur J Pharmacol 412:127-138.
- Alvarez-Fischer D, Blessmann G, Trosowski C, Béhé M, Schurrat T, Hartmann A, Behr TM, Oertel WH, Höglinger GU, Höffken H (2007) Quantitative [(123)I]FP-CIT pinhole SPECT imaging predicts striatal dopamine levels, but not number of nigral neurons in different mouse models of Parkinson's disease. Neuroimage 38:5-12.
- Niñerola-Baizán A, Rojas S, Bonastre M, Tudela R, Lomeña F, Pavía J, Marin C, Ros D (2015) In vivo evaluation of the dopaminergic neurotransmission system using [123I]FP-CIT SPECT in 6-OHDA lesioned rats. Contrast Media Mol Imaging 10:67-73.
- Marshall V, Grosset DG (2003) Role of dopamine transporter imaging in the diagnosis of atypical tremor disorders. Mov Disord 18 Suppl 7:S22-S27.
- Mateos JJ, Lomeña F, Parellada E, Font M, Fernandez E, Pavia J, Prats A, Pons F, Bernardo M (2005) Decreased striatal dopamine transporter binding assessed with [123I] FP-CIT in first-episode schizophrenic patients with and without shortterm antipsychotic-induced parkinsonism. Psychopharmacology (Berl) 181:401-406.
- Lavalaye J, Linszen DH, Booij J, Dingemans PM, Reneman L, Habraken JB, Gersons BP, van Royen EA (2001) Dopamine transporter density in young patients with schizophrenia assessed with [123]FP-CIT SPECT. Schizophr Res 47:59-67.
- Nikolaus S, Antke C, Kley K, Beu M, Wirrwar A, Müller HW (2009) Pretreatment with haloperidol reduces (123)I-FP-CIT binding to the dopamine transporter in the rat striatum: an in vivo imaging study with a dedicated small-animal SPECT camera. J Nucl Med 50:1147-1152.
- Booij J, van Loon G, de Bruin K, Voorn P (2014) Acute administration of haloperidol does not influence 123I-FP-CIT binding to the dopamine transporter. J Nucl Med 55:647-649.
- Seeman P (2002) Atypical antipsychotics: mechanism of action. Can J Psychiatry 47:27-38.
- Seeman P, Lee T, Chau-Wong M, Wong K (1976) Antipsychotic drug doses and neuroleptic/dopamine receptors. Nature 261:717-719.
- Schiffer WK, Mirrione MM, Biegon A, Alexoff DL, Patel V, Dewey SL (2006) Serial microPET measures of the metabolic reaction to a microdialysis probe implant. J Neurosci Methods 155:272-284.
- Paxinos G, Watson C (1986) The rat in stereotaxic coordinates. Academic Press, New York.
- Niñerola-Baizán A, Rojas S, Roé-Vellvé N, Lomeña F, Ros D, Pavía J (2015) Dopamine transporter imaging in the aged rat: a [123I]FP-CIT SPECT study. Nucl Med Biol 42:395-398.
- Fowler CJ, Benedetti MS (1983) The metabolism of dopamine by both forms of monoamine oxidase in the rat brain and its inhibition by cimoxatone. J Neurochem 40:1534-1541.
- Lane RF, Blaha CD (1987) Chronic haloperidol decreases dopamine release in striatum and nucleus accumbens in vivo: depolarization block as a possible mechanism of action. Brain Res Bull 18:135-138.
- Hernandez L, Hoebel BG (1989) Haloperidol given chronically decreases basal dopamine in the prefrontal cortex more than the striatum or nucleus accumbens as simultaneously measured by microdialysis. Brain Res Bull 22:763-769.
- Booij J, Andringa G, Rijks LJ, Vermeulen RJ, De Bruin K, Boer GJ, Janssen AG, Van Royen EA (1997) [123I]FP-CIT binds to the dopamine transporter as assessed by biodistribution studies in rats and SPECT studies in MPTP-lesioned monkeys. Synapse 27:183-190.
- Kim W, Park HS, Moon BS, Lee BC, Kim SE (2017) PET measurement of "GABA shift" in the rat brain: a preclinical application of bolus plus constant infusion paradigm of [18F] flumazenil. Nucl Med Biol 45:30-34.
- Basta-Kaim A, Budziszewska B, Jaworska-Feil L, Tetich M, Kubera M, Leśkiewicz M, Otczyk M, Lasoń W (2006) Antipsychotic drugs inhibit the human corticotropin-releasinghormone gene promoter activity in neuro-2A cells-an involvement of protein kinases. Neuropsychopharmacology 31:853-865.
- Sjøholm H, Bratlid T, Sundsfjord J (2004) 123I-beta-CIT SPECT demonstrates increased presynaptic dopamine transporter binding sites in basal ganglia in vivo in schizophrenia. Psychopharmacology (Berl) 173:27-31.
- Hsiao MC, Lin KJ, Liu CY, Tzen KY, Yen TC (2003) Dopamine transporter change in drug-naive schizophrenia: an imaging study with 99mTc-TRODAT-1. Schizophr Res 65:39-46.
- Laakso A, Bergman J, Haaparanta M, Vilkman H, Solin O, Syvälahti E, Hietala J (2001) Decreased striatal dopamine transporter binding in vivo in chronic schizophrenia. Schizophr Res 52:115-120.
- Schmitt GJ, Meisenzahl EM, Frodl T, La Fougère C, Hahn K, Möller HJ, Dresel S (2005) The striatal dopamine transporter in first-episode, drug-naive schizophrenic patients: evaluation by the new SPECT-ligand[99mTc]TRODAT-1. J Psychopharmacol 19:488-493.
- Yoder KK, Hutchins GD, Morris ED, Brashear A, Wang C, Shekhar A (2004) Dopamine transporter density in schizophrenic subjects with and without tardive dyskinesia. Schizophr Res 71:371-375.
- Yang YK, Yu L, Yeh TL, Chiu NT, Chen PS, Lee IH; SPECT study (2004) Associated alterations of striatal dopamine D2/ D3 receptor and transporter binding in drug-naive patients with schizophrenia: a dual-isotope SPECT study. Am J Psychiatry 161:1496-1498.
- Laakso A, Vilkman H, Alakare B, Haaparanta M, Bergman J, Solin O, Peurasaari J, Räkköläinen V, Syvälahti E, Hietala J (2000) Striatal dopamine transporter binding in neurolepticnaive patients with schizophrenia studied with positron emission tomography. Am J Psychiatry 157:269-271.
- Laruelle M, Abi-Dargham A, van Dyck C, Gil R, D'Souza DC, Krystal J, Seibyl J, Baldwin R, Innis R (2000) Dopamine and serotonin transporters in patients with schizophrenia: an imaging study with [(123)I]beta-CIT. Biol Psychiatr 47:371-379.
- Kasper S, Tauscher J, Willeit M, Stamenkovic M, Neumeister A, Küfferle B, Barnas C, Stastny J, Praschak-Rieder N, Pezawas L, de Zwaan M, Quiner S, Pirker W, Asenbaum S, Podreka I, Brücke T (2002) Receptor and transporter imaging studies in schizophrenia, depression, bulimia and Tourette's disorder--implications for psychopharmacology. World J Biol Psychiatry 3:133-146.
- Ham S, Kim TK, Chung S, Im HI (2017) Drug abuse and psychosis: new insights into drug-induced psychosis. Exp Neurobiol 26:11-24.