Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2019; 28(6): 658-669
Published online December 31, 2019
https://doi.org/10.5607/en.2019.28.6.658
© The Korean Society for Brain and Neural Sciences
EF-hand like Region in the N-terminus of Anoctamin 1 Modulates Channel Activity by Ca2+ and Voltage
Min Ho Tak1, Yongwoo Jang2, Woo Sung Son3, Young Duk Yang3* and Uhtaek Oh1,4*
1Interdisciplinary Program in Neuroscience, Seoul National University, Seoul 08826, 2Department of Biomedical Engineering, Hanyang University, Seoul 04763, 3College of Pharmacy, CHA University, Seongnam 13488, 4Sensory Research Center, Brain Science Institute, Korea Institute of Science & Technology (KIST), Seoul 02792, Korea
Correspondence to: *To whom correspondence should be addressed.
Young Duk Yang, TEL: 82-31-881-7170, FAX: 82-31-881-7219
e-mail: ntsky0816@cha.ac.kr
Uhtaek Oh, TEL: 82-2-958-7031, FAX: 82-2-958-7034
e-mail: utoh@kist.re.kr
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License
(http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and
reproduction in any medium, provided the original work is properly cited.
Abstract
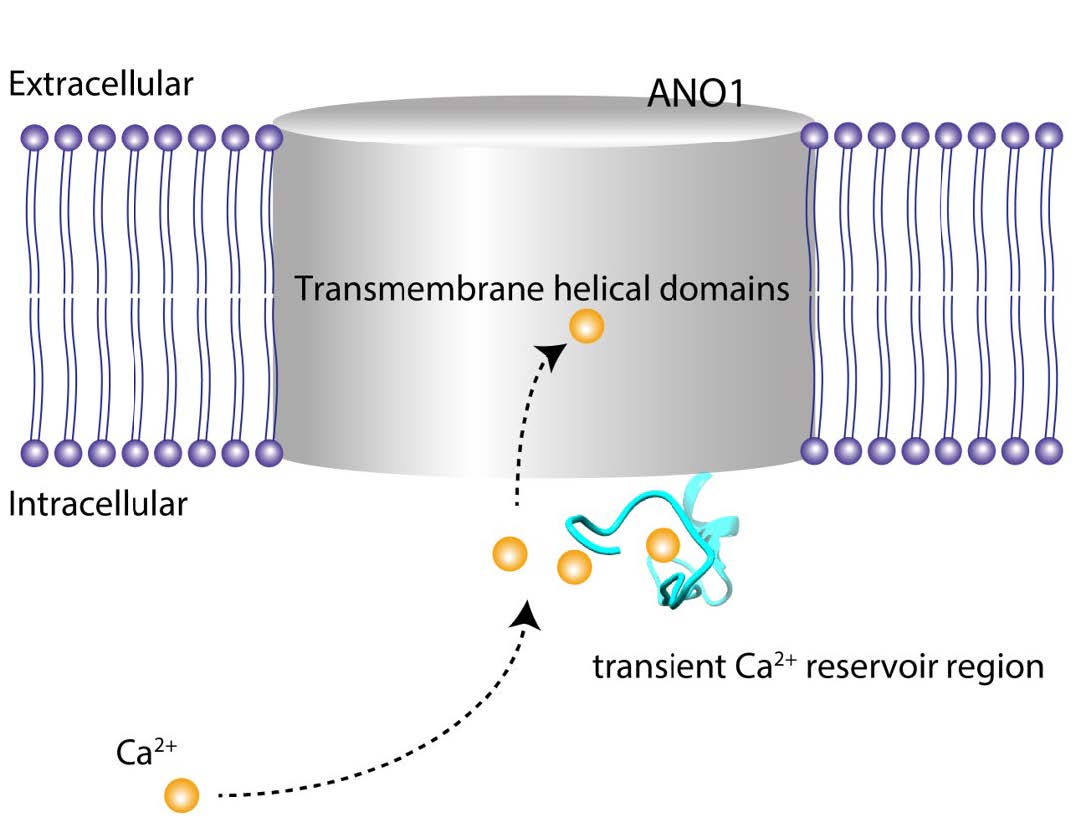
Anoctamin1 (ANO1) also known as TMEM16A is a transmembrane protein that functions as a Ca2+ activated chloride channel. Recently, the structure determination of a fungal Nectria haematococca TMEM16 (nhTMEM16) scramblase by X-ray crystallography and a mouse ANO1 by cryo-electron microscopy has provided the insight in molecular architecture underlying phospholipid scrambling and Ca2+ binding. Because the Ca2+ binding motif is embedded inside channel protein according to defined structure, it is still unclear how intracellular Ca2+ moves to its deep binding pocket effectively. Here we show that EF-hand like region containing multiple acidic amino acids at the N-terminus of ANO1 is a putative site regulating the activity of ANO1 by Ca2+ and voltage. The EF-hand like region of ANO1 is highly homologous to the canonical EF hand loop in calmodulin that contains acidic residues in key Ca2+-coordinating positions in the canonical EF hand. Indeed, deletion and Ala-substituted mutation of this region resulted in a significant reduction in the response to Ca2+ and changes in its key biophysical properties evoked by voltage pulses. Furthermore, only ANO1 and ANO2, and not the other TMEM16 isoforms, contain the EF-hand like region and are activated by Ca2+. Moreover, the molecular modeling analysis supports that EF-hand like region could play a key role during Ca2+ transfer. Therefore, these findings suggest that EF-hand like region in ANO1 coordinates with Ca2+ and modulate the activation by Ca2+ and voltage.
Graphical Abstract
Keywords: Anoctamin-1, Chloride channels, Calcium, Mutagenesis, Site-directed
INTRODUCTION
Endogenous Ca2+-activated chloride channels (CaCCs) regulate anionic currents by intracellular Ca2+ as well as by voltage [1–4]. Although CaCCs mediate diverse functions in various tissues, their biophysical properties display remarkable homogeneity. Their typical properties can be summarized as; activation by intracellular Ca2+ and voltage, an outwardly rectifying current-voltage relationship, slow time-dependent activation by voltage pulses, and voltage-dependent slow decay of tail currents [2]. Anoctamin 1 (ANO1) cloned for a CaCC has the biophysical and the pharmacological properties of native CaCCs [5–7]. For example, it is activated by Ca2+- and voltage, blocked by CaCC inhibitors, stimulated by G-protein-coupled receptor ligands via the PLC/IP3 pathway, expressed in transport epithelia, and has an outwardly rectifying I~V relationship [5–7]. ANO1 has four different splice variants, and of these, splice variants with exon 6b or exon 13 are known to affect Ca2+ sensitivity [8].
Because CaCCs are essential for mediating various physiological functions, an understanding of their activation mechanisms induced by intracellular Ca2+ and voltage would provide important insight of their functions. Several studies proposed the gating mechanism and putative binding residues by intracellular Ca2+. For example, Xiao and colleagues report that a poly-glutamate region in the first intracellular loop is important for coupling the voltage- and Ca2+-dependent gating of ANO1 [9]. Moreover, Lee and colleagues suggested that two helices in the third intracellular loop coordinate a conformational change in a Ca2+-dependent push-and-pull fashion [10]. Based on a positively charged ion of Ca2+, Tien and colleagues conducted systemic mutagenesis of all conserved acidic residues of ANO1, and concluded four putative amino acids (E650, E698, E701, and E730) are critical for Ca2+-induced ANO1 activation [11]. Recently, the crystal structure of the fungus
Although the Ca2+ binding site was determined in an nhTMEM16, the action mechanism underlying the activation of mammalian ANO1 and ANO2 by Ca2+ has still remained unclear. Based on the crystal structure of an nhTMEM16, the Ca2+ binding motif is deeply embedded inside phospholipid bilayer. Therefore, it is assumed the modulatory residues to help the movement of Ca2+ ions to the inside Ca2+ binding pocket of ANO1. In the present study, we set out to find the important residues to modulate channel activation of mouse ANO1. Especially, we investigated the role of the EF-hand like region in the N-terminus of ANO1 in both Ca2+- and voltage-dependent ANO1 activation.
MATERIALS AND METHODS
Mutagenesis and gene expression
All mutants were generated from the wild type construct, mouse ANO1 (pEGFP-N1-mANO1). Amino-acid substitution or deletion mutants were made using a site-directed mutagenesis kit (Muta-direct, iNtRON Biotech, Seoul, Korea) or by the overlap-PCR method. Mutations in all mutants were confirmed by sequencing whole nucleotide sequences.
HEK 293T cells were transfected with 1 μg of pEGFP-N1-mANO1 or pEGFP-N1-mutants, 0.05 μg pEGFP-N1, and FuGENE (Roche Diagnostics, Penzberg, Germany) in 35-mm Petri dishes. Transfected cells were incubated in DMEM supplemented with 10% fetal bovine serum (GIBCO) and penicillin-streptomycin at 37°C in a 5% CO2 incubator. Cells were used one or two days after plating.
Electrophysiology
As soon as a borosilicate glass pipette (World Precision Instruments, Sarasota, FL) touched the surface of a HEK cell, a gentle suction was applied to the pipette to obtain gigaseal. The tip resistance of the pipette was about 3 Mohms for whole-cell recordings. To record whole-cell currents, the membrane in contact with the pipette was ruptured by applying gentle suction after forming gigaseals. Once a whole cell was formed, the capacitive transients were canceled. Currents were recorded with a patch-clamp amplifier (Axopatch 200B, Molecular Devices) and filtered at 5 KHz. The whole-cell currents activated by each Ca2+ concentration (0.1~100 uM) were recorded from a holding potential of −60 mV during voltage clamp steps from −100 to+100 mV by 20 mV increments with 765 ms (−60 mV : 65 ms, −100~100 mV : 500 ms, −120 mV : 200 ms) total duration. Data from the amplifier were digitized with Digidata 1440A (Molecular Devices) and stored in a personal computer.
ANO1 currents were activated by intracellular Ca2+ at a holding potential of −60 mV unless specified. For recording currents in response to voltage pulses, membrane potential was changed from −100 mV to+100 mV in 20 mV increment. Square voltage pulses of 500 ms duration were delivered from a holding potential of −60 mV. Tail currents were recorded at −120 mV.
The control pipette solution contained 140 mM NMDG-Cl, 2 mM MgCl2, and 10 mM HEPES adjusted to pH 7.2. For the Ca2+–free solution, 1 mM EGTA was added to the control solution.
To calculate the accurate free intracellular Ca2+ concentration ([Ca2+]i) in pipette solutions, the WEBMAXC program in website (http://www.stanford.edu/~cpatton/webmaxc/webmaxcS.htm) was used. The bath solution contained 140 mM NMDG-Cl, 2 mM MgCl2, and 10 mM HEPES and was adjusted to pH 7.2.
Whole cell peak currents were recorded at each free [Ca2+]i, and then averaged for subsequent analysis. Half-maximal concentrations (EC50) were calculated using averaged peak currents, and fitted using the Hill equation. Deactivation time constants (τd) were derived by the single exponential fitting of tail currents recorded at −120 mV for 150 ms.
Molecular modeling of ANO1
To construct three-dimensional structure of mouse ANO1 using
RESULTS
Mutation in the EF-hand like region of ANO1 changes activation kinetics evoked by Ca2+
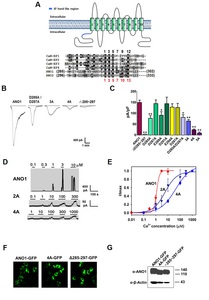
In mammals, calmodulin is a multifunctional intermediate protein that is regulated by Ca2+. The intracellular Ca2+ ions have well known to directly bind to EF-hand motif that is an signature sequence of the most common (canonical) EF-hand (-DxDx-DGxxxxxE-) of calmodulin [22, 23]. Interestingly, we first found that the amino acid residues (285-DGDYEGDNVE-294) in the N-terminus of ANO1 are somewhat similar to the EF-hand motif of calmodulin (Fig. 1A). Thus, we designated this region as the EF-hand like region in the present study. To investigate the effect of EF-hand like region on the activity of ANO1, we constructed ANO1 mutants in the region and then compared their activity by Ca2+ and voltage with those of the wild-type.
After forming whole cells with 10 μM Ca2+ in the pipette, large, robust currents were evoked in HEK cells transfected with wildtype ANO1. However, whole-cell currents to Ca2+ desensitized rapidly (Fig. 1B and 1C). In contrast to wild-type ANO1, whole-cell currents of Δ285~297 mutant to 10 μM Ca2+ were barely detectable (Fig. 1B and 1C). Therefore, Ala substitution was performed to determine the important amino acids in this region. More specifically, negatively charged amino acids, Asp or Glu at positions 285, 287, 289, or 291 were replaced by Ala. The Ala-substitution analysis revealed that D287A and D291A mutants elicited current responses comparable to that of wild type ANO1. However, the D285A and E289A mutants showed significant reductions in current response (Fig. 1B and 1C). The double (D285A/D287A) and triple Ala (D285A/D287A/E289A, 3A) substitution mutants also showed significant reductions in current density to 10 μM Ca2+ (Fig. 1B and 1C). When four or five acidic amino acids in 285-
To further investigate the responsiveness to Ca2+, different concentrations of Ca2+ were applied to the bath of inside-out membrane patches to investigate dose-response relationships. Because single-channel currents of ANO1 mutants activated by Ca2+ were too small to measure at −60 mV, the holding potential was set at +80 mV in order to increase current amplitudes. When the concentration of Ca2+ was increased from 0.01 μM to 10 μM, macroscopic currents were observed to increase dose-dependently (Fig. 1D and 1E). The effective concentration (EC50) of Ca2+ for the activation of wild-type ANO1 was 1.0 μM (n=5~11). Endogenous CaCCs are known to have a greater sensitivity to Ca2+ at depolarization [2–4]. Likewise, ANO1 also shows a slight increase in sensitivity to Ca2+ at depolarization. At −80 mV, the EC50 of ANO1 was 1.4 μM (n=5~12). Furthermore, the macroscopic channel currents of ANO1 increased abruptly between 0.3 and 3 μM. Thus, the Hill coefficient of wild-type ANO1 was found to be 3.7, suggesting a high level of cooperativity among Ca2+ coordination sites. In contrast, magnitudes of macroscopic currents of
Mutation in the EF-hand like region of ANO1 changes activation kinetics evoked by voltage
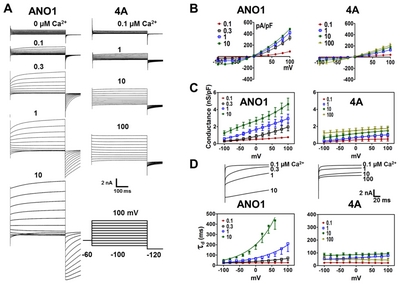
Voltage is also required for the activation of ANO1 [5]. However, the location in ANO1 responsible for its reaction to voltage pulses is not known. To examine ANO1 responses to voltage, voltage pulses from −100 mV to 100 mV were applied in 20 mV increments to HEK cells expressing the wild-type or
To investigate the kinetics of deactivation, tail currents activated by −120 mV after various pre-pulses were measured at different [Ca2+]i. As was observed for the tail currents of endogenous CaCCs [2, 4, 24], ANO1 tail currents decayed slowly as the pre-pulse depolarized (Fig. 2A and 2D), which suggests the voltage-dependent deactivation of ANO1. Furthermore, tail current decay was also found to be Ca2+ dependent, as tail currents decayed more slowly on increasing [Ca2+]i (Fig. 2D). The deactivation time constant (τd) was well fitted by a single exponential. τd increased as [Ca2+]i increased, especially at [Ca2+]i>1 μM. But at low [Ca2+]i (<0.3 μM), τd did not change appreciably as voltage was changed. Furthermore, hyperpolarization (−100 mV) deactivated ANO1 rapidly even in the presence of high [Ca2+]i (10 μM) (Fig. 2A and 2D).
In contrast, the
The EF-hand like region in ANO2
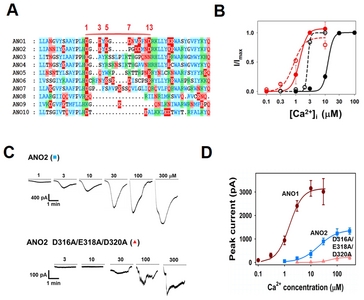
Of the 10 known isoforms of the anoctamin channel family, ANO1 and ANO2 are known to be activated by Ca2+ [5, 25]. Thus, we aligned the ~45 amino acids flanking the EF-hand like regions of all ANO homologs. As shown in Fig. 3A, acidic amino acids in 285-DGDYEGDNVEFND-297 of ANO1 were found to be well conserved in ANO2, but not in the other ANO homologs. This coincidence of homology and Ca2+ response also suggests that the EF-hand loop-like region is involved in the modulation for the Ca2+ sensitivity of ANO1 and ANO2.
ANO2 was found to be activated by [Ca2+]i in a dose-dependent manner (Fig. 3B). ANO2 also showed voltage-dependent Ca2+ sensitivity (Fig. 3C and 3D). At a holding potential of −80 mV, the EC50 for the Ca2+ activation of ANO2 was 12.5 μM. However, at +80 mV the EC50 reduced to 2.6 μM. Therefore, as observed in ANO1, ANO2 also showed a greater sensitivity to Ca2+ at depolarization than at hyperpolarization.
Because single-channel currents of ANO2 mutants were too small to measure even at the maximum Ca2+ concentration used (300 μM), whole-cell currents of wild-type ANO2 and of its D316A/E318A/D320A mutant were used to obtain an approximation of the dose-response relationships of ANO2 mutants (Ehold=−60 mV). The overall shape of ANO2 current response to Ca2+ was found to be similar to that of ANO1, except that a higher [Ca2+]i was required for activation. Appreciable currents were activated only by Ca2+ concentrations higher than 3 μM. The EC50 of ANO2 was found to be 20.0 μM, which is a 20-fold greater than that of ANO1 (EC50=1.0 μM) (Fig. 3C and 3D). When three acidic amino acids (316-DGEYDSPGDDMND-328) in the EF-hand like region in ANO2 were replaced with Ala (D316A/E318A/D320A), Ca2+ sensitivity and current density were substantially reduced (EC50=54.3 μM vs 20 μM for wild type ANO2) (Fig. 3C and 3D). These acidic amino acids correspond to the 1, 3, and 5 residues of the common EF hand of calmodulin, and are known to be essential for Ca2+ coordination [23]. These results further suggest that the acidic amino acids in the EF-hand like regions of ANO1 and ANO2 are an important modulatory residue in Ca2+-induced ANO1 activation.
Structural properties of ANO1 from molecular modeling analysis
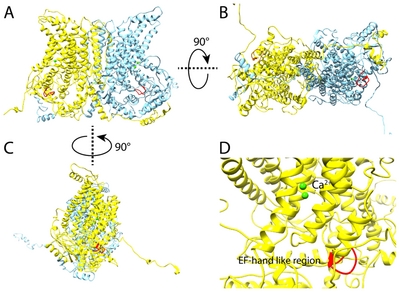
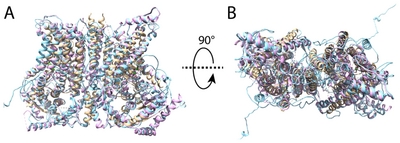
To ascertain the 3D structure-based involvement of EF-hand like region in Ca2+ binding to ANO1, we performed molecular modeling analysis using various in silico methods such as I-TASSER, ROSETTA [19], RaptorX [26], Phyre2 [27]. All modeled structures were validated using PSVS to select reasonable structure among predicted structures, and several modeled structures from I-TASSER algorithm was selected for further refinement. Finally, the best one structure among modeled structures was selected based on energy-scoring function and validation results. This model shows that the overall folding of ANO1 structure was similar with that of nhTMEM16 structure used as template during homology modeling (Fig. 4).
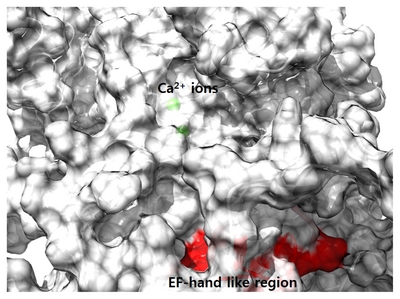
The three-dimensional structure of ANO1 shows a similar overall shape when compared to the nhTMEM16 structure used for modeling (Fig. 5). ANO1 has bundle of transmembrane helices like nhTMEM16, and specifically, the N-terminal region is longer than nhTMEM16. The long N-terminal region is composed of more than 120 amino acids and is expected to form a flexible loop rather than a transmembrane helix. Before reaching the first TM, several helix forms existed and some helical region seems to be able to bind Ca2+ ion. In modeled structure, the EF-hand like region was located at the entrance leading to the Ca2+ binding site (Fig. 6). Therefore, this EF-hand like region could have effect on the migration of Ca2+ ions between the cytoplasm and the Ca2+ binding site (Fig. 4 and 7).
DISCUSSION
Activation of ANO1 by Ca2+ and voltage
As is observed for native CaCCs [2, 4, 24], the rate of ANO1 activation by depolarization increases with [Ca2+]i. In addition, the deactivation of ANO1 was also found to be Ca2+ and voltage dependent. Kuruma et al. [2] derived a model for the Ca2+- and voltage-dependent activation and deactivation mechanisms of native CaCCs, which fits well with actual current responses to Ca2+ and voltage. This model contains three main features. First, the activations of CaCCs depend mainly on Ca2+, and not on voltage. Second, more than one Ca2+ ion is required to activate CaCCs, and third, deactivation depends on voltage. Evidently, ANO1 appears to possess all three of these features of endogenous CaCCs. First, ANO1 was easily activated by Ca2+ but rarely activated by voltage at low [Ca2+]i (Fig. 2A). Second, the Hill coefficient of Ca2+ for the activation of wild-type ANO1 was 3.7, and this reduced to less than 1 when Glu and Asp residues in the EF-hand like region were replaced by Ala. These results suggest that the binding of more than one Ca2+ is required for ANO1 activation and that acidic amino acids in the EF-hand like region cooperatively collect and transport Ca2+. Third, the deactivation of ANO1 is voltage dependent, because hyperpolarization was found to deactivate ANO1 even in the presence of Ca2+ (Fig. 2). Thus, the mechanisms underlying the activation and deactivation of ANO1 by Ca2+ and voltage appear to be similar to those proposed for native CaCCs.
Mechanism of Ca2+ binding and subsequent ion conduction of mANO1
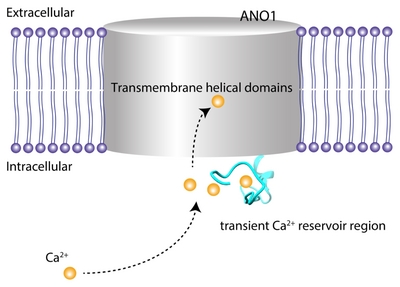
The Ca2+ binding of sites of ANO1 are well characterized in detail in cryo-EM studies [13, 28]. According to these studies, Cl− conducting pores are present in each subunit, which are formed by transmembrane helices α3-α7. The pore has a narrow neck that determines Cl− conductance and wide vestibule [13, 28]. When Ca2+ binds four acidic amino acids in α7 and α8, the positive charge density in α7 and α8 attracts the E654 residue in α6, which makes a comparably large movement of α6. This conformational change of α6 induces the opening of the neck of the pore, thus conducting Cl− [13]. Mammalian ANO1 has longer cytoplasmic N-terminal extension than nhTMEM16, whose function is not well characterized. The long region of N-terminal is located adjacent to the membrane surface rather than the region passing through the membrane. We do not know how the EF-hand like region in the N-terminus links to the Ca2+ binding sites in α6, α7, or α8. The strong phenotype of mutants in this region clearly suggests its role in modulating Ca2+-dependent activation of ANO1. Perhaps, the EF-hand like region acts like a transient Ca2+ reservoir, which could help the Ca2+ ions move to the Ca2+ binding site embedded inside the lipid bilayer (Fig. 7).
The EF-hand like region controls voltage dependence of ANO1
One of the canonical properties of CaCCs is its activation by Ca2+ and voltage [2]. As a CaCC, ANO1 is also activated by voltage and Ca2+ [5, 7, 9, 29]. The activation of ANO1 by voltage requires intracellular Ca2+ because without Ca2+, depolarization fails to open ANO1 [9, 29]. Thus, it seems the voltage- and Ca2+-dependent gating of ANO1 is tightly coupled. A few sites were suggested to be essential for the voltage- and Ca2+-dependent activation of ANO1. Structural analysis combined to mutagenesis study suggests that the sixth TM is essential for Ca2+ as well as voltage-dependent gating of mouse ANO1 [30]. A highly charged segment in the first intracellular loop (444–EEEEEAVK-451), an intracellular loop between TM α2 and TM α3, is important for Ca2+- and voltage-dependent gating [9, 29]. Deletion or mutation of EAVK residues profoundly reduced Ca2+ and voltage-activated currents suggesting the Ca2+ and voltage sensitivities are also coupled [9, 29]. The EF-hand like region in ANO1 appears to control the voltage dependence of ANO1. When we replaced four acidic amino acids in the EF-hand like region with Ala (the AGAYAGA mutant), the slow activation of ANO1 by voltage at low Ca2+, voltage-dependent conductance change, and voltage dependent increases in τd were largely eliminated (Fig. 2), which strongly suggest that the EF-hand like region also controls the voltage dependence of ANO1. As the voltage dependency relies on the binding of Ca2+ to the Ca2+ binding sites in the TM α6 whose conformational change gates the channel, it is conceivable that the ionic interaction between Ca2+ and oxygen atoms of carboxyl or carbonyl groups of the acidic amino acids in the EF-hand like region function as a voltage sensor. Voltage change in this region may affect the ionic interaction between Ca2+ and carboxyl or carbonyl groups in the region. Alternatively, the acidic amino acids in the EF-hand like region may interact with voltage sensors in the pore region or other areas as shown in the voltage-gated proton channel, Hv1 [31].
Structural element of the EF-hand like region of ANO1
EF-hand like regions in Ca2+ sensors and buffers usually has common structural features. In particular, canonical EF-hand regions commonly contain Asp and Glu residues in their 1, 3, 5, 9, and 12 positions for Ca2+ coordination [22, 23]. Furthermore, EF-hand proteins have a helix-loop-helix structure, and the helix-loop-helix motifs are present in pairs. In the present study, we performed computer modeling to determine the 3-dimensional structure of the EF-hand like region of ANO1. However, this region of ANO1 does not share similarity with the 3-dimensional structure of the EF hand of calmodulin, largely because helices flanking the loop region are lacking. Furthermore, the EF-hand like region in ANO1 is not present in pairs. Thus, this region in ANO1 does not fall into the canonical EF hand category. However, the primary structure of the ANO1 EF-hand like region is closely related to that of the EF hand because acidic amino acids are aligned well with those of the canonical EF-hand like region. For example, in canonical EF hands, Asp and Glu residues at the 1, 3, 5, 9, and 12 positions provide side-chain carboxylic oxygens for Ca2+ coordination [22, 23]. In addition, the backbone carbonyl oxygen at position 7 is also essential for Ca2+ chelation. Likewise, in the EF-hand like region of ANO1, Asp and Glu residues are present at the 1, 3, 5, 10, and 13 positions, which also presumably provide side-chain carboxylic oxygens for Ca2+ coordination. In addition, Glu at position 7 also provides a side-chain carboxylic oxygen for Ca2+ coordination. Thus, the structural element in this region of ANO1 explains why this region is essential for the activation by Ca2++ (page 20~21).
ACKNOWLEDGEMENTS
This research was supported by a grant from the National Research Foundation (NRF) of Korea (NRF-2018R1D-1A1B07049840) and the National Research Foundation of Korea, funded by the Ministry of Science and ICT (2011-0018358). Also, this work was partly supported by the GRRC program of Gyeonggi province (GRRC-CHA2017-A02, Validity and Safety Evaluation of Regional Specialized Resources).
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
- Hartzell C, Putzier I, Arreola J (2005) Calcium-activated chloride channels. Annu Rev Physiol 67:719-758.


- Kuruma A, Hartzell HC (2000) Bimodal control of a Ca(2+)activated Cl(-) channel by different Ca(2+) signals. J Gen Physiol 115:59-80.

- Arreola J, Melvin JE, Begenisich T (1996) Activation of calciumdependent chloride channels in rat parotid acinar cells. J Gen Physiol 108:35-47.
- Nilius B, Prenen J, Voets T, Van den Bremt K, Eggermont J, Droogmans G (1997) Kinetic and pharmacological properties of the calcium-activated chloride-current in macrovascular endothelial cells. Cell Calcium 22:53-63.
- Yang YD, Cho H, Koo JY, Tak MH, Cho Y, Shim WS, Park SP, Lee J, Lee B, Kim BM, Raouf R, Shin YK, Oh U (2008) TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 455:1210-1215.
- Caputo A, Caci E, Ferrera L, Pedemonte N, Barsanti C, Sondo E, Pfeffer U, Ravazzolo R, Zegarra-Moran O, Galietta LJ (2008) TMEM16A, a membrane protein associated with calciumdependent chloride channel activity. Science 322:590-594.
- Schroeder BC, Cheng T, Jan YN, Jan LY (2008) Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 134:1019-1029.
- Ferrera L, Caputo A, Ubby I, Bussani E, Zegarra-Moran O, Ravazzolo R, Pagani F, Galietta LJ (2009) Regulation of TMEM16A chloride channel properties by alternative splicing. J Biol Chem 284:33360-33368.
- Xiao Q, Yu K, Perez-Cornejo P, Cui Y, Arreola J, Hartzell HC (2011) Voltage- and calcium-dependent gating of TMEM16A/Ano1 chloride channels are physically coupled by the first intracellular loop. Proc Natl Acad Sci U S A 108:88918896.
- Lee J, Jung J, Tak MH, Wee J, Lee B, Jang Y, Chun H, Yang DJ, Yang YD, Park SH, Han BW, Hyun S, Yu J, Cho H, Hartzell HC, Oh U (2015) Two helices in the third intracellular loop determine anoctamin 1 (TMEM16A) activation by calcium. Pflugers Arch 467:1677-1687.
- Tien J, Peters CJ, Wong XM, Cheng T, Jan YN, Jan LY, Yang H (2014) A comprehensive search for calcium binding sites critical for TMEM16A calcium-activated chloride channel activity. Elife 3:e02772.
- Brunner JD, Lim NK, Schenck S, Duerst A, Dutzler R (2014) X-ray structure of a calcium-activated TMEM16 lipid scramblase. Nature 516:207-212.
- Paulino C, Neldner Y, Lam AK, Kalienkova V, Brunner JD, Schenck S, Dutzler R (2017) Structural basis for anion conduction in the calcium-activated chloride channel TMEM16A. Elife 6:e26232.
- Zhang Y (2008) I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 9:40.
- Pearlman DA, Case DA, Caldwell JW, Ross WS, Cheatham III TE, DeBolt S, Ferguson D, Seibel G, Kollman P (1995) AMBER, a package of computer programs for applying molecular mechanics, normal mode analysis, molecular dynamics and free energy calculations to simulate the structural and energetic properties of molecules. Computer Physics Communications 91:1-41.
- Case DA, Cheatham TE 3rd, Darden T, Gohlke H, Luo R, Merz KM Jr, Onufriev A, Simmerling C, Wang B, Woods RJ (2005) The Amber biomolecular simulation programs. J Comput Chem 26:1668-1688.
- Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE (2004) UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25:1605-1612.
- Meng EC, Pettersen EF, Couch GS, Huang CC, Ferrin TE (2006) Tools for integrated sequence-structure analysis with UCSF Chimera. BMC Bioinformatics 7:339.
- Bonneau R, Tsai J, Ruczinski I, Chivian D, Rohl C, Strauss CE, Baker D (2001) Rosetta in CASP4: progress in ab initio protein structure prediction. Proteins Suppl 5:119-126.
- Bhattacharya A, Tejero R, Montelione GT (2007) Evaluating protein structures determined by structural genomics consortia. Proteins 66:778-795.
- Konc J, Janežič D (2017) ProBiS tools (algorithm, database, and web servers) for predicting and modeling of biologically interesting proteins. Prog Biophys Mol Biol 128:24-32.
- Grabarek Z (2006) Structural basis for diversity of the EFhand calcium-binding proteins. J Mol Biol 359:509-525.
- Gifford JL, Walsh MP, Vogel HJ (2007) Structures and metalionbinding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem J 405:199-221.
- Evans MG, Marty A (1986) Calcium-dependent chloride currents in isolated cells from rat lacrimal glands. J Physiol 378:437-460.
- Ha GE, Cheong E (2017) Spike frequency adaptation in neurons of the central nervous system. Exp Neurobiol 26:179185.
- Wang S, Li W, Liu S, Xu J (2016) RaptorX-Property: a web server for protein structure property prediction. Nucleic Acids Res 44:W430-W435.
- Kelley LA, Mezulis S, Yates CM, Wass MN, Sternberg MJ (2015) The Phyre2 web portal for protein modeling, prediction and analysis. Nat Protoc 10:845-858.
- Dang S, Feng S, Tien J, Peters CJ, Bulkley D, Lolicato M, Zhao J, Zuberbühler K, Ye W, Qi L, Chen T, Craik CS, Jan YN, Minor DL Jr, Cheng Y, Jan LY (2017) Cryo-EM structures of the TMEM16A calcium-activated chloride channel. Nature 552:426-429.
- Xiao Q, Cui Y (2014) Acidic amino acids in the first intracellular loop contribute to voltage- and calcium- dependent gating of anoctamin1/TMEM16A. PLoS One 9(6): e99376.
- Peters CJ, Gilchrist JM, Tien J, Bethel NP, Qi L, Chen T, Wang L, Jan YN, Grabe M, Jan LY (2018) The sixth transmembrane segment is a major gating component of the TMEM16A calcium-activated chloride channel. Neuron 97:1063-1077. e4.
- DeCoursey TE (2018) Voltage and pH sensing by the voltagegated proton channel, HV1. J R Soc Interface 15:20180108.