Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Review Article
Exp Neurobiol 2017; 26(1): 1-10
Published online February 28, 2017
https://doi.org/10.5607/en.2017.26.1.1
© The Korean Society for Brain and Neural Sciences
Segregated Cell Populations Enable Distinct Parallel Encoding within the Radial Axis of the CA1 Pyramidal Layer
Tristan Geiller1,2, Sebastien Royer2 and June-Seek Choi1*
1Department of Psychology, Korea University, Seoul 02841, 2Center for Functional Connectomics, Korea Institute of Science and Technology, Seoul 02792, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-3290-2069
e-mail: j-schoi@korea.ac.kr
Abstract
Numerous studies have implicated the hippocampus in the encoding and storage of declarative and spatial memories. Several models have considered the hippocampus and its distinct subfields to contain homogeneous pyramidal cell populations. Yet, recent studies have led to a consensus that the dorso-ventral and proximo-distal axes have different connectivities and physiologies. The remaining deep-superficial axis of the pyramidal layer, however, remains relatively unexplored due to a lack of techniques that can record from neurons simultaneously at different depths. Recent advances in transgenic mice, two-photon imaging and dense multisite recording have revealed extensive disparities between the pyramidal cells located in the deep and the superficial layers. Here, we summarize differences between the two populations in terms of gene expression and connectivity with other intra-hippocampal subregions and local interneurons that underlie distinct learning processes and spatial representations. A unified picture will emerge to describe how such local segregations can increase the capacity of the hippocampus to compute and process numerous tasks in parallel.
Keywords: Hippocampus, CA1, pyramidal cell layer, Memory, place cell
INTRODUCTION
In the middle of the 20th century, the case of an amnesic patient shed light on the role of the hippocampus in forming and organizing newly acquired memory of facts and events, namely declarative memory. Following a surgical ablation of his medial temporal lobe, Henry Molaison, better known as patient HM, had his procedural and working memory preserved but was not capable of remembering facts and events that had just occurred if recalled later than 30 min [1,2]. These observations were among the first to link memory functions to a specific part of the brain and ignited burgeoning research toward identifying the role and anatomical components of what later had been called the medial temporal lobe memory system [3,4].
Later, the discovery of place-modulated activity in hippocampal pyramidal cells (PC) of rats foraging in a box raised new questions on the neural substrate of spatial navigation and orientation [5,6]. Those findings brought the hippocampus to the fore, a structure that had been mainly studied as a model for long-term potentiation of synaptic transmission thanks to its orderly segregation into layers [7]. Indeed, the hippocampus is traditionally divided into four functional regions, namely the dentate gyrus (DG), and three subdivisions of the Cornu Ammonis (CA) area, on the basis of anatomical criteria [8,9,10], connectivity [11] and functional features [12,13]. During the last few decades, studies have focused on building models exposing regional differences in memory encoding or navigation [14,15,16,17,18,19,20].
The four regions and their interconnections form the hippocampal circuit. The full wiring map is complicated, but the general tendency has been to start with the entorhinal cortex (EC) and build around a sequential excitatory circuit. In this model, the EC projects to the DG through the perforant path, then the granule cells of the DG contact CA3 through the mossy fibers. These subfields are believed to be an important area where integrated stimulus from the cortex are separated and sorted to retrieve previously encoded memory with minimal interference [20,21]. Next, the CA3 pyramidal cells project via the Schaffer collaterals to CA1, considered an integration area where high-level computation is executed to achieve cognitive processes [22,23,24,25]. Finally, CA1 outputs to subiculum, back to the entorhinal cortex, and related parahippocampal regions.
This simplified model became more complex with the growing number of research on functional hippocampal architecture. New wiring diagrams now accommodate recent anatomical and physiological data and notably include the subfield CA2 as well as direct projections from the EC to the CA1 region.
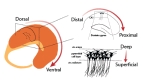
Beyond these differences among the main subregions, recent evidences indicated functional segregations along the principal axes of the hippocampus. The dorso-ventral axis has perhaps drawn the first attention on potential differences as suggested by the anatomical asymmetries of the hippocampus (Fig. 1) [27,28]. First, the dorsal and the ventral parts do not make connections to the same brain area, as the dorsal part targets mainly the septal pole and the ventral part the temporal pole [29,30]. In addition to these connectivity patterns, the two parts further differentiate by distinct gene expression. Lastly, the role of the dorsal hippocampus is believed to be computing cognitive functions while the ventral area relates to emotion and stress [31,32].
The second axis, also called transverse or proximo-distal axis, also exhibits gradual anatomical and physiological changes in both the CA1 and CA3 areas (Fig. 1). In CA1, the principal cell layer extends from the subiculum to the border of CA2. The proximal part (bordering CA2) receives more direct inputs from the medial entorhinal cortex (MEC) than the distal part (close to the subiculum) that in contrast is innervated by the lateral entorhinal cortex (LEC) [27]. The MEC nests the grid cells, a class of spatially modulated cells that fire in a grid-like pattern when the animal forages in an open arena, whereas the LEC conveys information relative to non-spatial objects [33,34,35]. Henriksen et al. [36] have shown distinct spatial representations that gradually varied along this proximo-distal axis. Place fields in the proximal part were more precise and spatially tuned than their distal counterparts, partly due to the strong space-modulated nature of the MEC inputs they receive.
Such functionally transverse organization was also found in CA3 where both the Knierim's and Moser's teams found concordant dissimilarities between the proximal and distal part of the pyramidal layer [22,37]. Their major findings showed that the exposition of the animal to different environment triggered a greater remapping in proximal CA3 area than in distal. This suggests a functional dissociation with proximal CA3 being notably involved in pattern separation.
Despite the flourishing number of studies on dorso-ventral and proximo-distal contrasts, the deep-superficial axis of the pyramidal cell layer has just recently started to present evidence of a radially organized circuitry (Fig. 1). These late findings are partly due to the uniform anatomical aspects of pyramidal cells that made it nearly impossible to distinguish potential heterogeneities, although the functional diversification of interneurons has been relatively well documented [38]. Furthermore, the lack of recording techniques to precisely capture the verticality has long prevented from revealing differences along the depth of small local areas.
The purpose of the current review is to bring together the latest findings on functional differences within the CA1 radial axis of the pyramidal cell layer and to propose theoretical and computational implications of such local organization. First, we will describe the morphological and genetic diversity that resides within the layer, and discuss how those differences may explain the distinct connectivity between the deep and superficial part. The last section will focus on the laminated electrophysiological features responsible for dissimilar encoding of space and memory.
MORPHOLOGY AND MOLECULAR DIFFERENCES
The pyramidal cells in CA1 do not visually segregate in two distinct layers per se, but are distributed uniformly with a gradient of distinct morphological, molecular and physiological features along the radial axis. As the terminology is borrowed from cortical anatomy, the definition of deep and superficial part of the layer is based on the location of the neurons' basal versus apical dendrites. Consequently, superficial CA1 (sCA1), bordering
The
Morphologically, it was found in primate monkeys that the arborization of the apical dendrites were different for cell bodies depending on their depth in the layer [44]. Indeed, the cells in the superficial part showed a more extensive apical dendritic arborization that supports a higher number of terminal's apical branches than in the deep part. A result that was not seen in rodent studies. This significance of this observation is questionable but was not seen in rodent studies and therefore may translate in neural processing differences between the two species.
Heterogeneities can also be found during early developmental stages where the earlier-born neurons position in the upper row of the CA1 pyramidal layer, and later-born neurons in the lower row [45,46]. Taking advantage of this distinct neurogenesis, Cembrowski et al. [47] performed labeling
At the level of the cell membrane, a recent study revealed that cannabinoid receptors and hyperpolarization-activated cyclic nucleotide-gated (HCN) channels were involved in the control of learning and memory, but that their activation differed as a function of the cell layer's depth [51]. On one hand, the cannabinoid type-1 receptors (CB1R) can modulate the release of excitatory or inhibitory neurotransmitter upon their activation [52,53]. On the other hand, HCN channels mediate the hyperpolarization-activated cationic depolarizing current (Ih), believed to exert a stabilizing effect on neuronal excitability by their negative-feedback properties on controlling the resting potential of the cellular membrane [54]. Both CB1R and Ih currents have globally been designated responsible for a wide range of cognitive functions and synaptic mechanisms in the hippocampus [55,56,57,58], but had not yet been seen to cell specific. The study by Maroso et al. first presented that the application of CB1R agonist increased the postsynaptic Ih only in superficial pyramidal cells as shown by an increase in the hyperpolarization-induced depolarizing sag. Although both deep and superficial cells exhibited Ih currents upon membrane hyperpolarization, no increase was seen in deep pyramidal neurons. In addition to electrophysiological data, they performed an object location memory (OLM) task to test whether this CB1R-Ih pathway was involved in spatial memory formation. Following the exploration of an environment enriched with two objects, a mouse was treated with CB1R agonist or saline and reexposed 24 h later to the same environment where one of the object was moved. Interestingly, the activation of the CB1R-Ih pathway in only superficial cells was sufficient to strongly alter spatial memory and reduce the time spent by the mice in exploring the moved object against the trained object. The work by Maroso et al. was the first demonstration that the CB1R-Ih pathway functioned exclusively in superficial cells of the CA1 layer, and that it served mnemonic purposes as assessed by the OLM test.
CONNECTIVITY IN THE HIPPOCAMPAL CIRCUIT
Here, we will review the latest findings in terms of afferent and efferent connections to distinct cell population within the CA1 pyramidal layer, as well as their contribution to perform segregated yet interlaced deep versus superficial local computations. Slomianka et al. [59] have reviewed and summarized studies in regards of efferent connectivity, but a lack of studies prevented the characterization of different inputs projecting onto either the deep or superficial CA1 PCs. Therefore, the following part will focus on discussing these aspects.
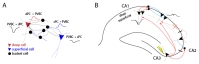
Lorente de Nó [10] was first to predict a non-uniform connectivity of interneurons onto principal cells in the hippocampus but it is only 80 years later that Lee et al. [60] provided evidence of such a local microcircuit mediating inhibition and excitation. Parvalbumin (PV)-expressing interneurons have been largely studied and identified to play multiple functions in several brain areas [61]. In the hippocampus, they appear to be crucial in the generation of the 8~12 Hz theta rhythm [62]. More precisely in CA1, PV+ interneurons have various morphologies that underlie distinct functional properties and target-specificity (e.g somatic versus dendritic inhibition) [38]. Lee et al. focused on the perisomatic GABAergic inhibition provided by Parvalbumin-expressing basket cells (PVBC) onto the deep or superficial CA1 PCs. Performing whole-cell recordings, they observed physiological differences in response to intracellular current injected. Superficial pyramidal cells (sPC) showed a more depolarized resting potential as well as larger sag potentials in response to hyperpolarizing current pulses, consistent with data reported by Maroso et al. and described in the previous section. Simultaneous paired recordings of PVBC and PC interestingly revealed a heterogeneous innervation of PC by PVBC that resulted in larger inhibitory post-synaptic currents (IPSC) in deep pyramidal cells (dPC) (Fig. 2A). These bigger postsynaptic currents were mediated by PVBC's perisomatic axon terminals that were found in larger number on dPCs than on sPCs (Fig. 2A). Accordingly, the innervation of PVBC by PC was not homogeneous either as superficial cells had a higher probability of excitatory connections onto PVBC interneurons (Fig. 2A). However, the probability of glutamate release and the amplitude of excitatory postsynaptic current (EPSC) in PVBC was not different whether the stimulated pyramidal cell was located in the superficial or deep part of the layer (Fig. 2A). This study revealed a new form of excitatory-inhibitory loop within the principal cells and local interneurons, where sPCs are more likely to target PVBCs that in turn control dPCs' spiking. While some studies had already attributed specific roles to interneuron subtypes in generation of rhythm or spiking in the hippocampus [63], it would be of value to reexamine these findings taking into account the uneven connectivity of interneurons with deep and superficial pyramidal cells as indicated by the work of Lee et al.
At a larger scale, the Tonegawa group found a new interregional pathway stemming from the dentate gyrus, projecting to CA2 pyramidal cells that in turn synapse onto deep CA1 [64]. Combining cell-specific transgenic mice and immunochemistry, Hogawa et al. showed that CA2 pyramidal cells project to CA1 in a uniform manner by sending their axons evenly into the
These recent discoveries of interregional and local circuitry involving the deep and superficial pyramidal cells in CA1 shed light on the complex dynamic of excitation and inhibition during sharp-wave-ripple (SWR) events [65] (Fig. 2B). SWR is a high-frequency (150~250 Hz) oscillatory event in the hippocampus thought to be critical for memory encoding and consolidation. However, it is still unclear how ripples are initiated and whether it involves an oscillatory response between distinct classes of cells. Stark et al. [43] first pointed out a time difference in the spiking activity during SWR cycles. Extracellular recordings of thousands of cells with multi-site silicon probes revealed that sPCs fired earlier and at a higher probability than their deep peers. In their study using intracellular recordings, Valero et al. [65] revealed the complex and synchronized circuit that dominates during SWR. Upon the electrical activation of CA3 fibers, the excitation propagates to downstream regions of CA1 and CA2 (Fig. 2B). Pyramidal cells in CA2 are de-inhibited, and therefore limit the excitation given to dPCs in CA1, consistent with the results described previously [64] (Fig. 2B). In addition, dPCs remain inactive due to the large inhibitory control from PVBC, as shown in Lee et al. Furthermore, they found that sPCs were more prominently contacted by another class of interneurons, the Cholecystokinin (CCK)-containing cells, whose sparse firing pattern leaves sPCs under the excitation provided by CA3 fibers (Fig. 2B). This excitation and the higher probability of sPCs to contact PVBC accentuates the inhibitory drive onto dPCs (Fig. 2B). Overall, these findings prove that SWR events regulate a finely tuned local circuit within the hippocampus that results in a gradient of activity radially distributed along the CA1 pyramidal layer. Although the hypothesis has not been tested yet, these distinct deep-superficial dynamics during SWR may underlie more advanced computational functions. For instance, it still unknown where and how events are partitioned and filtered out before being sent into long-term memory storage.
FUNCTIONAL SEGREGATION ALONG THE DEEP AND SUPERFICIAL LAYERS IN SPATIAL NAVIGATION AND LEARNING
The development of new techniques for large-scale recordings of neuronal activity has opened the door to a much finer and precise characterization of ensembles and dynamics in various parts of the brain. The range of benefits is broad and considerable. For instance, Mizuseki et al. [66] exploited the geometry of the silicon probes to first sample a superficial-deep subdivision of hippocampal cells as rats foraged in freely moving conditions. Their study showed gradual differences of electrophysiological features along the deep-superficial axis of the pyramidal layer in rats. More precisely, they showed that sPCs tended to have less place fields than dPCs in an open arena or a linear track where rats foraged for food pellets. They also confirmed
Mizuseki et al. also looked at how the cells reorganized their spiking activity depending on the behavioral state of the animal. For this purpose, they successfully monitored the same pyramidal cells during both awake foraging and sleep periods after the animals returned to their home cage. During sleep, the phase relationship of the neurons' spikes to the local field potential gradually shifted with the location of the neuron along the pyramidal layer, with the superficial part showing no change and the deep part showing the biggest shift. The local field potential is always carefully examined in the hippocampus as rhythms are thought to conduct synchronization of cell ensembles and enable a time frame for sequential activity [69]. Specific rhythms can also reflect the communication of two brain structures [70,71]. The most prominent is the theta rhythm (8~12 Hz) found notably during rapid eye-movement (REM) sleep and awake exploration [72]. In the study of Mizuseki et al., deep pyramidal cells shifted the theta phase at which they preferentially discharge during foraging by 180 degrees during REM sleep, and were consequently labeled REM-shifting cells. Taking this analysis further, they also showed that REM-shifting and non-REM-shifting cells had a different preferred gamma (30~100 Hz) phase during awake epochs. These findings are interesting as the phase-dependent spiking activity is thought to reveal potential connectivity across different brain structures. Therefore, it indicates that layer-specific inputs projecting onto the hippocampus potentially dominate in a brain state-dependent manner.
Danielson et al. [73] performed the first optical demonstration by simultaneously imaging the calcium activity of both superficial and deep CA1 cells thanks to a 2-photon microscope coupled with a piezoelectric crystal that allowed them to quasi-instantly switch the field of view between the two layers. Imaging data confirmed electrophysiology data on the higher probability of deep cells to exhibit place fields during behavioral tasks. They monitored both layers while mice were engaged either in a random foraging task or a goal-oriented learning (GOL) task adapted for their treadmill apparatus. During random foraging, they observed that superficial cells were less subject to a remapping, defined as the change in spiking activity following a change of the context (i.e change of belt and cues on the treadmill). sPCs were also more stable after the repeated exposure of the same context across days. On the contrary, deep pyramidal cells displayed a higher stability across sessions in a task that required the animals to learn the position of a hidden reward. The assessment of a day-by-day fields' stability showed that dPCs tended to gradually change their spiking location toward the neighborhood of the reward while the superficial cells did not show such progressive transformation. According to Danielson et al. findings, the neural substrate of learning resides in the deep layer of the CA1 pyramidal cells while the superficial layer is involved in spatial navigation and foraging.
This further confirms the idea of a dual stream of information conveyed by distinct pathways consistent with Hogawa et al.'s findings [64]. Further experiments could tell if a layer-specific inactivation would impair learning or spatial memory although the idea that these complex processes reside in such a small and local population of cells remains questionable.
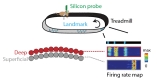
Finally, the study by Geiller et al. [74] showed segregated place field mechanisms along the deep-superficial axis of the CA1 pyramidal cell layer underlying different spatial representations. Taking advantage of the linearity that a treadmill offers, they trained mice to run on a belt onto which several different landmarks were attached (Fig. 3, top). The mice performed a simple foraging task as the reward was kept at the same position across all the sessions and trials. Following dense recording with silicon probes, they found that a portion of cells located in the deep layer of CA1 had their firing field tightly correlated with the location of landmarks of the same nature (Fig. 3). On the other hand, the superficial layer contained cells that exhibited mostly single firing fields, less tied to the landmarks' location and thus more likely to represent a more general context as a whole (Fig. 3). Those results are consistent with previous work reporting the influence of landmark in a vector-dependent manner on CA1 principal cells (LV cells) [75]. This previous study, however, could not estimate the depth of the recorded cells within the layer, as tetrodes did not allow the spatial resolution along the Z-axis. Therefore, they could not distinguish whether LV cells and non-LV cells formed distinct cell populations. Geiller et al. study reveals that a subset of cells located in the deep portion of the pyramidal layer is more likely to be linked with environmental sensory stimulus while the superficial portion has more flexible but global coding of space that may originate from the CA3 region. This demonstration once again favors the argument of a parallel processing of information that may underlie distinct cognitive functions.
CONCLUSION
The hippocampus has been the subject of many hypotheses, but its precise role in memory, navigation and learning is yet to be determined [76]. In fact, the hippocampus is associated with multiple brain functions but no single model addresses the issue of how it manages to accomplish these various tasks [77,78]. For instance, there are parallel lines of research looking into memory encoding and spatial navigation. Although the two views still diverge, novel findings support a bridging framework to reconcile space and memory and answer the question of how distinctively different information is processed within the same structure [79,80].
Until very recently, hippocampus' models and theories were built on a view of homogenous population of principal cells, regardless of their spatial distribution along different axes. Coding and computations were thought to be handled individually by the main CA regions, and information to follow a unidirectional pathway confined in excitatory monosynaptic connections from the dentate gyrus to CA3, then from CA3 to CA1. Only a small but growing number of literature highlighted segregated connections between the dorsal and ventral hippocampus, respectively projecting to brain areas responsible for cognitive functions (prefrontal cortex) and emotions (amygdala). Within the dorsal hippocampus, it then became the turn of the proximo-distal axis to be in the spotlight with findings revealing uneven projections originating from the medial and lateral area of the entorhinal cortex and targeting the CA1 region. Yet, and despite old anatomical reports, it seemed unlikely to observe local differences residing in the superficial-deep axis of pyramidal cells akin to cortical lamination.
However, morphological, gene expression and neuronal birthdate differences all indicated an eventual disparity between the pyramidal cells contained in the deep part versus the superficial part of CA1. The study on GABAergic interneurons confirmed the possibility that excitation and inhibition was not equally balanced but rather mediated by a local laminated microcircuit. In spite of these findings, it was still unclear if two or more clearly distinct cell populations dwelled together but formed functional different units. Electrophysiology results using silicon probes and 2-photon imaging to record simultaneously deep and superficial cells of rodents engaged in behavioral tasks revealed such functional layer-specificities. On the one hand, deep CA1 cells showed stability upon learning, had more place fields during navigation though more subject to remapping upon repeated exposure of the same environment, and controlled by brain state-dependent inputs. Considering the more bursty nature of those deep pyramidal cells, one would hypothesize that they are more subject to rapid transformation through synaptic plasticity and therefore are the substrate for flexible encoding. On the other hand, superficial cells were more active during memory-related ripple events, and are thought to encode spatial contexts as a whole. Their less adjustable spiking activity makes them a robust candidate for short-term memory depository before being sent or changed to a long-term memory. Altogether, these new findings bring on several implications for the dissection of the microcircuits regulating computations in the hippocampus, beyond a simple organization within the pyramidal layer. The superficial-deep specificities combined with dorsal-ventral and proximo-distal disparities add another level of complexity, or one more degree of freedom for possible distinct information's pathways.
Figures
{kind=link}
{kind=link}
{kind=link}
References
- Corkin S. Lasting consequences of bilateral medial temporal lobectomy: clinical course and experimental findings in H.M. Semin Neurol 1984;4:249-259.
- Salat DH, van der Kouwe AJ, Tuch DS, Quinn BT, Fischl B, Dale AM, Corkin S. Neuroimaging H.M.: a 10-year follow-up examination. Hippocampus 2006;16:936-945.

- Mishkin M. Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature 1978;273:297-298.
- Squire LR. The legacy of patient H.M. for neuroscience. Neuron 2009;61:6-9.
- O'Keefe J, Dostrovsky J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res 1971;34:171-175.
- O'Keefe J, Nadel L. The hippocampus as a cognitive map. Oxford: Clarendon Press, 1978.
- Bliss TV, Lomo T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J Physiol 1973;232:331-356.
- Golgi C. Sulla fina anatomia degli organi centrali del sistema nervosa. Milano: Hoepli, 1886.
- Ramón y Cajal S, Pasik P, Pasik T. Texture of the nervous system of man and the vertebrates: an annotated and edited translation of the original Spanish text with the additions of the French version by Pedro Pasik and Tauba Pasik. Wien: Springer, 1999.
- Lorente de Nó R. Studies on the structure of the cerebral cortex. Leipzig: Johann Ambrosius Barth, 1934.
- Andersen P, Bliss TV, Skrede KK. Lamellar organization of hippocampal pathways. Exp Brain Res 1971;13:222-238.
- Blackstad TW. Commissural connections of the hippocampal region in the rat, with special reference to their mode of termination. J Comp Neurol 1956;105:417-537.
- Amaral DG. Emerging principles of intrinsic hippocampal organization. Curr Opin Neurobiol 1993;3:225-229.
- Teyler TJ, DiScenna P. The role of hippocampus in memory: a hypothesis. Neurosci Biobehav Rev 1985;9:377-389.
- McNaughton BL, Barnes CA, Gerrard JL, Gothard K, Jung MW, Knierim JJ, Kudrimoti H, Qin Y, Skaggs WE, Suster M, Weaver KL. Deciphering the hippocampal polyglot: the hippocampus as a path integration system. J Exp Biol 1996;199:173-185.
- Leutgeb S, Leutgeb JK, Barnes CA, Moser EI, McNaughton BL, Moser MB. Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science 2005;309:619-623.
- Eichenbaum H, Dudchenko P, Wood E, Shapiro M, Tanila H. The hippocampus, memory, and place cells: is it spatial memory or a memory space?. Neuron 1999;23:209-226.
- Amaral DG, Witter MP. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience 1989;31:571-591.
- Lee I, Kesner RP. Differential contributions of dorsal hippocampal subregions to memory acquisition and retrieval in contextual fear-conditioning. Hippocampus 2004;14:301-310.
- Lee I, Yoganarasimha D, Rao G, Knierim JJ. Comparison of population coherence of place cells in hippocampal subfields CA1 and CA3. Nature 2004;430:456-459.
- Rolls ET. A quantitative theory of the functions of the hippocampal CA3 network in memory. Front Cell Neurosci 2013;7:98.
- Neunuebel JP, Knierim JJ. CA3 retrieves coherent representations from degraded input: direct evidence for CA3 pattern completion and dentate gyrus pattern separation. Neuron 2014;81:416-427.
- Andersen P. The hippocampus book. Oxford: Oxford University Press, 2007.
- Chen J, Olsen RK, Preston AR, Glover GH, Wagner AD. Associative retrieval processes in the human medial temporal lobe: hippocampal retrieval success and CA1 mismatch detection. Learn Mem 2011;18:523-528.
- Duncan K, Ketz N, Inati SJ, Davachi L. Evidence for area CA1 as a match/mismatch detector: a high-resolution fMRI study of the human hippocampus. Hippocampus 2012;22:389-398.
- Schlichting ML, Zeithamova D, Preston AR. CA1 subfield contributions to memory integration and inference. Hippocampus 2014;24:1248-1260.
- Paxinos G. The rat nervous system. San Diego: Academic Press, 1995.
- Nadel L. Dorsal and ventral hippocampal lesions of behavior. Physiol Behav 1968;3:891-900.
- Risold PY, Swanson LW. Structural evidence for functional domains in the rat hippocampus. Science 1996;272:1484-1486.
- Dolorfo CL, Amaral DG. Entorhinal cortex of the rat: topographic organization of the cells of origin of the perforant path projection to the dentate gyrus. J Comp Neurol 1998;398:25-48.
- Petrovich GD, Canteras NS, Swanson LW. Combinatorial amygdalar inputs to hippocampal domains and hypothalamic behavior systems. Brain Res Brain Res Rev 2001;38:247-289.
- Fanselow MS, Dong HW. Are the dorsal and ventral hippocampus functionally distinct structures?. Neuron 2010;65:7-19.
- Hafting T, Fyhn M, Molden S, Moser MB, Moser EI. Microstructure of a spatial map in the entorhinal cortex. Nature 2005;436:801-806.
- Knierim JJ, Neunuebel JP, Deshmukh SS. Functional correlates of the lateral and medial entorhinal cortex: objects, path integration and local-global reference frames. Philos Trans R Soc Lond B Biol Sci 2013;369:20130369.
- Tsao A, Moser MB, Moser EI. Traces of experience in the lateral entorhinal cortex. Curr Biol 2013;23:399-405.
- Henriksen EJ, Colgin LL, Barnes CA, Witter MP, Moser MB, Moser EI. Spatial representation along the proximodistal axis of CA1. Neuron 2010;68:127-137.
- Lu L, Igarashi KM, Witter MP, Moser EI, Moser MB. Topography of place maps along the CA3-to-CA2 axis of the hippocampus. Neuron 2015;87:1078-1092.
- Klausberger T, Somogyi P. Neuronal diversity and temporal dynamics: the unity of hippocampal circuit operations. Science 2008;321:53-57.
- Stephan H. Comparative anatomy of allocortex. Verh Anat Ges 1976;:217-251.
- Bennett MV, Pereda A. Pyramid power: principal cells of the hippocampus unite!. Brain Cell Biol 2006;35:5-11.
- Mercer A, Bannister AP, Thomson AM. Electrical coupling between pyramidal cells in adult cortical regions. Brain Cell Biol 2006;35:13-27.
- Pais I, Hormuzdi SG, Monyer H, Traub RD, Wood IC, Buhl EH, Whittington MA, LeBeau FE. Sharp wave-like activity in the hippocampus in vitro in mice lacking the gap junction protein connexin 36. J Neurophysiol 2003;89:2046-2054.
- Stark E, Roux L, Eichler R, Senzai Y, Royer S, Buzsáki G. Pyramidal cell-interneuron interactions underlie hippocampal ripple oscillations. Neuron 2014;83:467-480.
- Altemus KL, Lavenex P, Ishizuka N, Amaral DG. Morphological characteristics and electrophysiological properties of CA1 pyramidal neurons in macaque monkeys. Neuroscience 2005;136:741-756.
- Angevine JB, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature 1961;192:766-768.
- Caviness VS. Time of neuron origin in the hippocampus and dentate gyrus of normal and reeler mutant mice: an autoradiographic analysis. J Comp Neurol 1973;151:113-120.
- Cembrowski MS, Bachman JL, Wang L, Sugino K, Shields BC, Spruston N. Spatial gene-expression gradients underlie prominent heterogeneity of CA1 pyramidal neurons. Neuron 2016;89:351-368.
- Baimbridge KG, Peet MJ, McLennan H, Church J. Bursting response to current-evoked depolarization in rat CA1 pyramidal neurons is correlated with lucifer yellow dye coupling but not with the presence of calbindin-D28k. Synapse 1991;7:269-277.
- Dong HW, Swanson LW, Chen L, Fanselow MS, Toga AW. Genomic-anatomic evidence for distinct functional domains in hippocampal field CA1. Proc Natl Acad Sci U S A 2009;106:11794-11799.
- Nielsen JV, Blom JB, Noraberg J, Jensen NA. Zbtb20-induced CA1 pyramidal neuron development and area enlargement in the cerebral midline cortex of mice. Cereb Cortex 2010;20:1904-1914.
- Maroso M, Szabo GG, Kim HK, Alexander A, Bui AD, Lee SH, Lutz B, Soltesz I. Cannabinoid control of learning and memory through HCN channels. Neuron 2016;89:1059-1073.
- Katona I, Sperlágh B, Sík A, Käfalvi A, Vizi ES, Mackie K, Freund TF. Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J Neurosci 1999;19:4544-4558.
- Mackie K. Distribution of cannabinoid receptors in the central and peripheral nervous system. Handb Exp Pharmacol 2005;:299-325.
- Biel M, Wahl-Schott C, Michalakis S, Zong X. Hyperpolarization-activated cation channels: from genes to function. Physiol Rev 2009;89:847-885.
- Castillo PE, Younts TJ, Chávez AE, Hashimotodani Y. Endocannabinoid signaling and synaptic function. Neuron 2012;76:70-81.
- Nolan MF, Malleret G, Dudman JT, Buhl DL, Santoro B, Gibbs E, Vronskaya S, Buzsáki G, Siegelbaum SA, Kandel ER, Morozov A. A behavioral role for dendritic integration: HCN1 channels constrain spatial memory and plasticity at inputs to distal dendrites of CA1 pyramidal neurons. Cell 2004;119:719-732.
- Shah MM. Cortical HCN channels: function, trafficking and plasticity. J Physiol 2014;592:2711-2719.
- Soltesz I, Alger BE, Kano M, Lee SH, Lovinger DM, Ohno-Shosaku T, Watanabe M. Weeding out bad waves: towards selective cannabinoid circuit control in epilepsy. Nat Rev Neurosci 2015;16:264-277.
- Slomianka L, Amrein I, Knuesel I, Sørensen JC, Wolfer DP. Hippocampal pyramidal cells: the reemergence of cortical lamination. Brain Struct Funct 2011;216:301-317.
- Lee SH, Marchionni I, Bezaire M, Varga C, Danielson N, Lovett-Barron M, Losonczy A, Soltesz I. Parvalbumin-positive basket cells differentiate among hippocampal pyramidal cells. Neuron 2014;82:1129-1144.
- Hu H, Gan J, Jonas P. Interneurons. Fast-spiking, parvalbumin(+) GABAergic interneurons: from cellular design to microcircuit function. Science 2014;345:1255263.
- Amilhon B, Huh CY, Manseau F, Ducharme G, Nichol H, Adamantidis A, Williams S. Parvalbumin interneurons of hippocampus tune population activity at theta frequency. Neuron 2015;86:1277-1289.
- Royer S, Zemelman BV, Losonczy A, Kim J, Chance F, Magee JC, Buzsáki G. Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nat Neurosci 2012;15:769-775.
- Kohara K, Pignatelli M, Rivest AJ, Jung HY, Kitamura T, Suh J, Frank D, Kajikawa K, Mise N, Obata Y, Wickersham IR, Tonegawa S. Cell type-specific genetic and optogenetic tools reveal hippocampal CA2 circuits. Nat Neurosci 2014;17:269-279.
- Valero M, Cid E, Averkin RG, Aguilar J, Sanchez-Aguilera A, Viney TJ, Gomez-Dominguez D, Bellistri E, de la Prida LM. Determinants of different deep and superficial CA1 pyramidal cell dynamics during sharp-wave ripples. Nat Neurosci 2015;18:1281-1290.
- Mizuseki K, Diba K, Pastalkova E, Buzsáki G. Hippocampal CA1 pyramidal cells form functionally distinct sublayers. Nat Neurosci 2011;14:1174-1181.
- Jarsky T, Mady R, Kennedy B, Spruston N. Distribution of bursting neurons in the CA1 region and the subiculum of the rat hippocampus. J Comp Neurol 2008;506:535-547.
- Grosmark AD, Buzsáki G. Diversity in neural firing dynamics supports both rigid and learned hippocampal sequences. Science 2016;351:1440-1443.
- Buzsáki G, Anastassiou CA, Koch C. The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes. Nat Rev Neurosci 2012;13:407-420.
- Colgin LL, Denninger T, Fyhn M, Hafting T, Bonnevie T, Jensen O, Moser MB, Moser EI. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009;462:353-357.
- Benchenane K, Peyrache A, Khamassi M, Tierney PL, Gioanni Y, Battaglia FP, Wiener SI. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron 2010;66:921-936.
- Buzsáki G. Rhythms of the brain. Oxford: Oxford University Press, 2006.
- Danielson NB, Zaremba JD, Kaifosh P, Bowler J, Ladow M, Losonczy A. Sublayer-specific coding dynamics during spatial navigation and learning in hippocampal area CA1. Neuron 2016;91:652-665.
- Geiller T, Fattahi M, Choi JS, Royer S. Place cells are more strongly tied to landmarks in deep than in superficial CA1. Nat Commun 2017;8:14531.
- Deshmukh SS, Knierim JJ. Influence of local objects on hippocampal representations: Landmark vectors and memory. Hippocampus 2013;23:253-267.
- Buzsáki G, Moser EI. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci 2013;16:130-138.
- Milivojevic B, Doeller CF. Mnemonic networks in the hippocampal formation: from spatial maps to temporal and conceptual codes. J Exp Psychol Gen 2013;142:1231-1241.
- Maguire EA, Mullally SL. The hippocampus: a manifesto for change. J Exp Psychol Gen 2013;142:1180-1189.
- Eichenbaum H, Cohen NJ. Can we reconcile the declarative memory and spatial navigation views on hippocampal function?. Neuron 2014;83:764-770.
- Schiller D, Eichenbaum H, Buffalo EA, Davachi L, Foster DJ, Leutgeb S, Ranganath C. Memory and space: towards an understanding of the cognitive map. J Neurosci 2015;35:13904-13911.