Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2018; 27(3): 189-199
Published online June 30, 2018
https://doi.org/10.5607/en.2018.27.3.189
© The Korean Society for Brain and Neural Sciences
Antidepressant-like Effects of p-Coumaric Acid on LPS-induced Depressive and Inflammatory Changes in Rats
Seok Lee1†, Hyun-Bum Kim1†, Eun-Sang Hwang2, Eun-seok Kim3, Sung-Soo Kim3, Tae-Dong Jeon3, Min-cheol Song1, Ji-Seung Lee1, Min-Chan Chung3, Sungho Maeng2* and Ji-Ho Park1,4*
1Department of East-West Medical Science, Graduate School of East-West Medical Science, Kyung Hee University, Yongin 17104, 2Department of Gerontology, Graduate School of East-West Medical Science, Kyung Hee University, Yongin 17104, 3Department of East-West Medicine, Graduate School of East-West Medical Science, Kyung Hee University, Yongin 17104, 4Research Institute of Medical Nutrition, Kyung Hee University, Yongin 17104, Korea
Correspondence to: *To whom correspondence should be addressed.
Sungho Maeng, TEL: 82-31-201-2916, FAX: 82-31-204-8119
e-mail: jethrot@khu.ac.kr
Ji-Ho Park, TEL: 82-31-201-2180, FAX: 82-31-204-8119
e-mail: jihopark@khu.ac.kr
†These authors contributed equally
Abstract
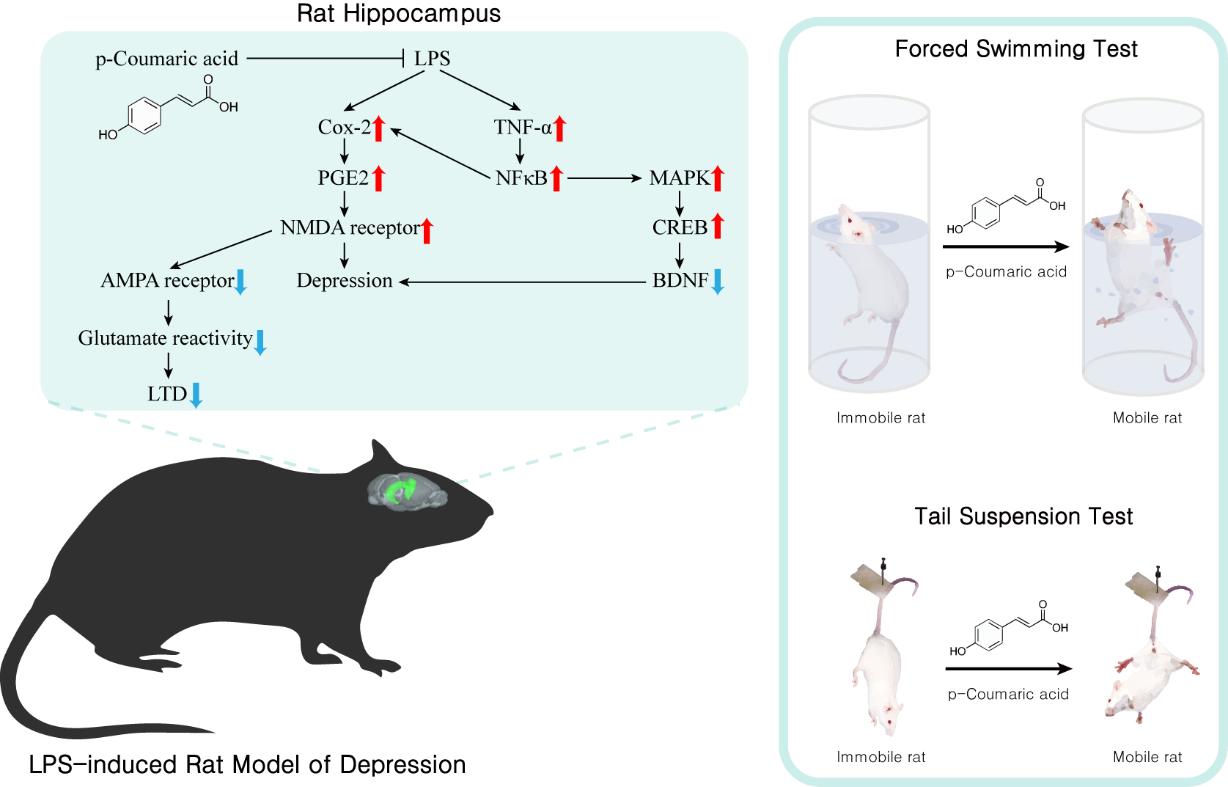
Depression causes mental and physical changes which affect quality of life. It is estimated to become the second most prevalent disease, but despite its commonness, the pathophysiology of depression remains unclear and medicine is not sufficiently protective. p-Coumaric acid (p-CA) is a dietary phenolic acid which has been proven to have antifungal, anti-HIV, anti-melanogenic, antioxidant and anti-inflammatory effects. Considering these effects, we investigated whether p-CA can prevent depressive symptoms by reducing inflammatory cytokines in animals injected with lipopolysaccharide (LPS). Changes in despair-related behaviors, inflammatory cytokines, neurotrophic factors and synaptic activity were measured. In these animals, p-CA improved despair-related behavioral symptoms induced by LPS in the forced swim test (FST), tail suspension test (TST) and sucrose splash test (SST). p-CA also prevented the increase of inflammatory cytokines in the hippocampus such as cycloxigenase-2 and tumor necrosis factor-α due to LPS. Similarly, it prevented the reduction of brain-derived neurotrophic factor (BDNF) by LPS. Electrophysiologically, p-CA blocked the reduction of long-term depression in LPS-treated organotypic tissue slices. In conclusion, p-CA prevented LPS-induced depressive symptoms in animals, as determined by behavioral, biochemical and electrophysiological measures. These findings suggest the potential use of p-CA as a preventive and therapeutic medicine for depression.
Graphical Abstract
Keywords: Depression, p-coumaric acid (p-CA), Lipopolysaccharide (LPS), Long-term synaptic depression (LTD), Cytokines
INTRODUCTION
Depression is an emotional disorder that causes mental and physical changes characterized by lack of motivation and melancholic feeling. These symptoms can affect the daily lives of the patients and can hardly be stopped by personal effort [1]. Recently, depression became one of the most prevalent health risks and the World Health Organization estimated that it will become the second-most common disease in 2020 [2]. Although it is becoming a more serious issue, the pathophysiology of depression still remains unclear [3].
Neurochemical research has improved our understanding of neurotransmitter changes in depression, which has contributed to the development of antidepressants such as TCAs (tricyclic antidepressants), SSRIs (selective serotonin reuptake inhibitors) and NDRIs (norepinephrine-dopamine reuptake inhibitors). However, the efficacy of these antidepressants is sometimes suboptimal and delayed in onset, which is problematic when depressed patients have suicidal ideation or are at risk of self-harm [4]. In addition, the limits of pharmacotherapy and pharmacological classification based on serotonin, norepinephrine and dopamine imply that the ‘monoamine hypothesis’ is woefully insufficient in explaining the etiology of depression. Interestingly, the inflammatory pathway has recently become an important topic in the development of novel antidepressants [5]. Therefore, prophylactic antidepressants with fewer side effects and more comprehensive effects should be developed [6].
Continuous inflammation with elevated levels of inflammatory cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) can cause sickness behavior that replicates depressive symptoms [7,8]. Depressed patients and animal models of depression show high levels of IL-6 and TNF-α in their cerebrospinal fluid and frontal cortex [9]. Such findings suggest that inflammatory cytokines are important contributors to the development of depression [10]. In support of this hypothesis, preclinical studies demonstrated that induction of cytokines using LPS or deletion of interleukin-1 (IL-1) [11], IL-6 [12] and TNF-α [13] can cause alterations in behavior, neurotransmitter function, neurogenesis and neuroendocrine output [14]. These inflammatory cytokines cause activation of glial cell and modify the monoamine neurotransmitters. Inflammatory cytokines activate serotonergic system and affect metabolism of 5-HT or NE [15]. These modifying of inhibitory and excitatory neurotransmitters affect emotional activity including NE or GABA, and these results impair the balance between neurotransmitters [16] which is directly linked to the depression.
In preclinical and translational research on depression, animal models have been developed to screen the core symptoms of depression by validating the endophenotypes of depression, such as learned helplessness [17] and anhedonia [18]. By screening these symptoms through behavioral tests, the severity of depression can be measured. Long-term depression (LTD) also has a potentially important role in modern neuroscience, and it is possible that they may be exploited to treat disorder and disease in the central nervous system. Previous studies have shown that LTD may be involved in depression and may be a useful tool to study depression. The molecular mechanisms underlying LTD could potentially rectify synaptic malfunction and lead to a treatment for depression. For instance, socially stressed rats demonstrated significant LTD in hippocampus 30 min after low-frequency stimulation while control groups didn't demonstrate LTD-induction. And, antidepressant such as imipramine blocked LTD in stressed group [19]. The molecular mechanisms underlying LTD could potentially rectify synaptic malfunction and lead to a treatment for depression.
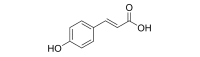
p-Coumaric acid (p-CA) is a phenolic compound (Fig. 1) commonly present in vegetables and fruits. It is used to make other flavonoids [20] and has been proven to have antifungal, anti-viral, anti-melanogenic, antioxidant and anti-inflammatory effects [21]. p-CA also has been reported to have neuroprotective effects equal to or greater than those of flavonoids (+)-catenchin, (−)-epicatechin and quercetin [22]. In practice, previous studies have shown that treatment with p-CA significantly diminished brain oxidative stress, decreased inflammation and inhibited apoptosis in the hippocampus [23]. These previous studies suggest that p-CA may function as an antidepressant because of its anti-inflammatory effects and neuroprotective effects.
Considering these beneficial effects, this study was performed to investigate whether the protective effects of p-CA can alleviate depressive symptoms involved with inflammatory cytokines. We used LPS to stimulate the release of pro-inflammatory cytokines which is known to induce depression-like behavior lasting for a few hours or even longer [10]. The protective effect of p-CA was then measured by behavioral, biochemical and electrophysiological methods.
MATERIALS AND METHODS
Thirty 6-week-old male Sprague-Dawley rats with an average weight of 200±20 g were used for
p-CA, LPS from Escherichia coli (L2637), HEPES (H4034), L-glutamine (G-8540) and D-glucose (G-7528) were purchased from Sigma (St. Louis, MO, USA). Minimum essential medium (MEM, LM 007-01), Hank's balanced salt solution (HBSS, LB 003-01) and horse serum (S 104-01) were purchased from JBI (Daegu, South Korea). Penicillin and streptomycin were obtained from Gibco BRL (LS 202-02, USA).
p-CA was dispersed in ethanol so that its final concentration was 20%. Rats were randomly assigned to six groups (n=6/group). Group 1 was the EtOH 20% group and received EtOH 20% for two days. Group 2 was the control group and received saline for two days. Group 3, the LPS group, received saline on the first day and LPS on the second day. Group 4 was given 50 mg/kg p-CA [24] on the first day and LPS on the second day. Group 5 was given 75 mg/kg p-CA [25] on the first day and LPS on the second day. Group 6 was given 100 mg/kg p-CA on the first day and LPS on the second day. The p-CA and EtOH was administered at a dose of 1mL per 100 g body weight. All drug administration was blinded. The p-CA and saline treatments were administered intraperitoneally between 10:00 a.m. and 11:00 a.m. a day before the behavioral experiments. LPS was dissolved in saline and injected at a dose of 200 µg/kg 2 h before initiating the behavioral experiments (Fig. 2) [10].
The FST was conducted in a sound-attenuated room to measure behavioral despair. The test was performed in two sessions as described previously, a pre-swim trial and the forced swim test, with a few changes [26]. The pre-swim trials and FST were performed in cylinders (diameter 24.4 cm) filled with tap water (23℃±2) to a depth of about 30 cm (Fig. 2). A white barrier was placed between the cylinders for visual isolation between animals. The pre-swim trial was 15 minutes and the FST was 24 hours after the pre-swim trial. After each trial, the animals were wiped with a towel and returned to their home cage, after which the cylinder was refilled with fresh water [4]. During the six-minute FST, all behavior was recorded to measure their despair behaviors. Immobility was defined as the sum of the time the animals remained floating without limb movement.
The TST is another behavioral test for assessing antidepressant activity through the display of despair and hopelessness. All behaviors were recorded during tail suspension to measure immobility. The rats were suspended 50 cm above the floor for 6 min by adhesive tape attached 3 cm from the tip of the tail. Rats were considered immobile only when they hung passively and were completely motionless [27].
The splash test is a pharmacologically validated animal model to exhibit motivational behavior. Decrease of grooming time in this test is considered an apathy-like behavior of depression [28]. The test was performed by squirting 10% sucrose solution on the dorsal coat of animals in their home cage. The time spent grooming was recorded for 5 minutes.
Rats were anesthetized with ketamine and hippocampus was collected. In brief, the hippocampus was denatured in lysis buffer with a tissue grinder. Then the homogenate was resolved by SDS-PAGE and transferred to a PVDF membrane. Primary antibodies (anti-COX-2, anti-TNF-α, anti-BDNF; Cell signaling Tech. Inc., Danvers, MA) were diluted to 1:1000 concentration ratio and applied overnight at 4℃. After a one-hour incubation in 1:1000 diluted secondary antibody, bands were visualized by enhanced chemiluminescence (ECL) solution (Thermo Fisher Scientific, MA) and quantified using a chemiluminescence detector (Davinch Chemi imaging system, CellTagen, Korea) and ImageJ (NIH, USA) image analysis software.
Seven-day-old Sprague-Dawley rats were sacrificed and their brains were quickly extracted and immersed in ice-cold HBSS medium (LB 003-01, Sigma, St. Louis, Mo, USA) with 20 mM HEPES (H-4034, Sigma, St. Louis, Mo, USA) immediately. The cerebellum and frontal cortex were removed delicately and the hippocampus was extracted and amputated transversely at 350 µm by a tissue chopper (Mickle Laboratory Engineering Co., Surrey, UK). In each six-well plate, four or five slices were placed onto a 0.4 µm culture insert (Millicell-CM; Millipore, Bedford, MA, USA). The six-well plate was filled with 1mL of culture medium (MEM; 25%(v/v) Hank's balanced salt solution, supplemented with 20 mM HEPES, 6 g/L D-glucose, 1 mM L-glutamine, 25%(v/v) heat-inactivated horse serum and 1% penicillin-streptomycin; pH=7.1). The culture was incubated at 36℃ with 5% CO2 and 95% O2. The culture medium was replaced three times a week and the sections were cultured for two weeks [4].
A single stabilized hippocampal slice was removed from a membrane insert delicately with a needle and then placed on an 8×8 microelectrode array (MEA) with 10-µm-diameter electrodes. They were spaced 100 µm apart (Multi Channel Systems GmbH, Reutlingen, Germany) and pre coated with 0.01% polyethylenimine. The slice was stabilized in artificial cerebrospinal fluid (aCSF), which includes 10 mM glucose, 114 mM NaCl, 3 mM KCl, 26 mM NaHCO3, 2 mM CaCl2, 1 mM MgCl2 and 20 mM HEPES, for 30 minutes at 33℃ and pH7.4 with 95% O2 and 5% CO2 gas aeration. Then, extraneous aCSF was extracted by a pipette. The MEA with the hippocampal slice was moved to the interface of an MEA1060 amplifier. The array solution was grounded by Ag/Ag pellets and data were collected from every channel with a speed of 25 kHz. They were recorded using Recorder-Rack software (MEA systems, MCS software). After the experiment, the array was cleansed with 2% Ultrasonol 7 (Carl Roth GmbH, Karlsruhe, Germany) in distilled water for 30 minutes, rinsed with distilled water and immersed in distilled water at room temperature.
The MEA system consisted of a 64-channel array containing 60 recording electrodes (STG1004; Multi Channel Systems GmbH, Reutlingen, Germany) and four stimulating electrodes, an amplifier (MEA1060; Multi Channel Systems GmbH, Reutlingen, Germany) in a Faraday cage, data acquisition software (www.multichannelsystems.com) and temperature control units (Multi Channel Systems GmbH, Reutlingen, Germany) [4]. Bipolar electrical stimulation was applied in the CA2 stratum radiatum region to stimulate the Schaffer collateral (SC) and commissural pathways. The intensity of the bipolar test pulse (or baseline) stimulation set was 100 mA, optimized to provide about 40~65% of the maximum tissue response, delivered once per minute. Before the low-frequency stimulation (1 Hz for 15 minutes; 900 total pulses) was applied to induce LTD, baseline responses were evoked for at least 30 minutes, where the last 40 minutes were recorded. From the 59 microelectrodes spanning the hippocampus, fEPSPs were recorded every 60 sec for another 50 minutes after the conditioning stimulation. During the experiment, the slices were continually perfused with new aCSF or aCSF in drugs (dissolved with 95% O2, 5% CO2) at 3 mL/min and 161/h carbogen consumption.
MC_Rack (v.3.2.1.0, MultiChannel Systems) was used to digitize the MEA signal and protect EPSPs from inducing amplitudes over 40 mV. Then, a custom MATLAB (v.7.0.1, Mathworks, Inc.) program was used to remove artificial factors and integrate the evoked field potential trajectory [29].
The results in the figures are expressed as mean±standard error of the mean (SEM). Statistical analysis was performed using one-way and repeated measure ANOVA followed by Scheffe or Dunnett T3 post-hoc multiple comparison test using SPSS 20.0 for Windows (SPSS Inc., Chicago, IL, USA). Statistically significant differences were defined as p<0.05.
RESULTS
Depression-like changes induced by LPS and the protective effect of p-CA were tested with animal models of behavioral despair and apathy. There were significant differences in immobility time and climbing time measured in the FST between groups (Fig. 3B and 3C). The one-way ANOVA Scheffe post-hoc test showed significant differences in accumulated immobility time for control vs. LPS (*p<0.05), LPS vs. p-CA 50 (#p<0.05) and LPS vs. p-CA 75 (##p<0.01), but the immobility reduction in the p-CA 100 group was insignificant (LPS vs p-CA 100). The ANOVA Scheffe post-hoc test showed significant differences in climbing time for control vs. LPS (**p<0.01), and LPS vs. p-CA 50 (##p<0.01), but not for LPS vs. p-CA 75 or LPS vs. p-CA 100.
There were also significant differences between groups in the immobility time measured in the TST (Fig. 3E). The one-way ANOVA Scheffe post-hoc test showed significant differences when comparing control vs. LPS (***p<0.001), LPS vs. p-CA 50 (##p<0.01), LPS vs. p-CA 75 (##p<0.01) and LPS vs. p-CA 100 (#p<0.05).
The grooming time in the SST was also significantly different between groups (Fig. 3F). A post-hoc pairwise comparison showed significant differences for control vs. LPS (**p<0.01), LPS vs. p-CA 50 (###p<0.001), LPS vs. p-CA 75 (##p<0.01) and LPS vs. p-CA 100 (##p<0.01). These results indicate that p-CA can improve LPS-induced depression-like behaviors.
Because p-CA was diluted in EtOH 20% as a vehicle, we compared the effect of EtOH 20% with saline treated animals. There was no difference between the two groups in the effect of saline and 20% ethanol (data not shown).
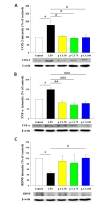
Hippocampal expression of cyclooxygenase-2 (COX-2), TNF-α and brain-derived neurotrophic factor (BDNF) was measured by immunoblotting (Fig. 4). LPS increased the expression of COX-2 (*p<0.05) and TNF-α (*p<0.05) but reduced the expression of BDNF (**p<0.01). When p-CA was co-treated with LPS, protein levels of TNF-α and COX-2 decreased significantly compared to animals treated with LPS only in a dose range of 50 to 100mg/kg (#p<0.05). Co-treatment of p-CA and LPS increased the expression of BDNF when p-CA was given at 100 mg/kg (#p<0.05) but was non-significantly increased at 50 mg/kg and 75 mg/kg. These results indicate that p-CA can prevent LPS-induced inflammatory changes and improve synaptic plasticity in the hippocampus.
The effects of LPS and p-CA were analyzed by comparing the average field excitatory postsynaptic potentials (fEPSPs) to record LTD in the hippocampus for 105 minutes (Fig. 5). The total fEPSP activity calculated from the experimental results showed a significant within-group effect, between-group effect (***p<0.001) and within-between group interaction. The Scheffe post-hoc test showed a significant reduction of total fEPSP in LPS-treated tissues (control vs. LPS: **p<0.01). p-CA had no effect on vehicle-treated tissues (control vs. p-CA: p=0.99) but improved fEPSP activity in LPS-treated tissues (LPS vs. LPS+p-CA: ##p<0.01) (Fig. 5A). The average fEPSP was also significantly different between groups (Fig. 5B). The Scheffe post-hoc test showed that LPS had a significant effect on the average fEPSP (control vs. LPS: **p<0.01). Similarly, p-CA had no effect on average fEPSP in the vehicle-treated group but improved the LPS-induced change (LPS vs LPS+p-CA: p<0.01). In conclusion, p-CA improved the LPS-induced changes in LTD in hippocampal tissues.
DISCUSSION
Depression involves many biological factors such as cytokines, glucocorticoids and monoamines [30]. At the molecular level, dysregulation of neurotransmitters, hormones, signaling pathways, neurotrophic and neuroplastic molecules and inflammatory mediators contribute to the pathophysiology of depression. In this study, LPS-treated rats were used to elucidate the extent to which brain inflammation contributes to depressive behaviors such as motivation reduction and learned helplessness, as well as how p-CA interferes with such behavior and molecular stress-induced alternations [31].
In the FST, which measures behavioral despair, LPS induced depression-like behaviors, thus increasing the immobility time. However, when p-CA was co-administered with LPS, depression-like behavior was significantly reduced, as shown by decreased immobility time and increased climbing time. In addition, immobility in TST, another measurement of behavioral despair, was increased by LPS, and p-CA also improved this depression-like phenotype in these animals. Moreover, grooming time in the SST was reduced by LPS treatment only but increased under co-treatment with p-CA, meaning that LPS and p-CA affected apathy, another indicator of depression [28]. Although not reported in p-CA yet, p-CA derivatives such as naringenin [32] and resveratrol [33] have been demonstrated to have antidepressant-like effects. For example, acute naringenin treatment reduced immobility time in the tail suspension test by altering the monoaminergic system [34] and affected sucrose preference in stressed animals by upregulating the hippocampal expression of BDNF [32]. Resveratrol demonstrated antidepressant-like effects by regulating the hypothalamus-pituitary-adrenal axis in Wistar-Kyoto rats, a putative and non-induced animal model of depression [35]. Climbing behavior was defined as a characteristic of norepinephrine reuptake blockers [26]. Fluoxetine selectively increased swimming in the FST, whereas the norepinephrine reuptake blocker, desipramine, only increased climbing. Because climbing increased for animals treated with a low dose of p-CA (50 mg/kg) but not for those treated with high doses (75 and 100 mg/kg). Reduced motivation is another component of depression, which is modeled by measuring grooming behaviors in the sucrose splash test. Imipramine and fluoxetine improved motivational behaviors in the splash test [36]. Ketamine also improved motivational behavior in socially defeated animals [37]. Our results demonstrate that p-CA improved motivational behaviors reduced by LPS. In conclusion, p-CA significantly improved the core symptoms of depression. These previous studies and our results suggest that p-CA is an anti-depressant. In this study, the climbing time was increased by p-CA in the low dose (50 mg/kg) while it didn't show significant effect in high doses (75 and 100 mg/kg). Also, the other results such as grooming time in SST and immobility in TST showed slightly more effective in low dose (50 mg/kg), which means low dose of p-CA showed more anti-depressant effect consistently in behavioral tests. These results suggest that p-CA probably regulates the noradrenergic and serotonergic system with different potency.
Next, the state of the neurons and the degree of depression-related inflammation was evaluated by measuring the hippocampal expression of inflammatory cytokines and a neurotrophic factor. Patients with major depression exhibited elevated levels of inflammatory cytokines in peripheral blood and brain tissues, which can be involved in the pathophysiology of depression through neurotransmitter metabolism, neural plasticity and neurotrophic support [38]. Meta-analysis has shown a significant correlation between increased TNF-α and IL-6 and depression [39]. Also, COX-2 inhibitor has been shown to improve depressive symptoms in both preclinical and clinical settings [40,41]. While both were increased by LPS, p-CA treatment reduced LPS-induced changes in COX-2 and TNF-α. Interestingly, depressed patients with increased inflammatory cytokine levels are more likely to become resistant to antidepressant therapies [42]. These reduced inflammation-related molecular changes demonstrate the potential of p-CA as an alternative for treatment-resistant depression. In this study, p-CA reduced inflammatory cytokines such as TNF-α and COX-2 in this study. COX-2 is increased by NFκB which is activated by TLR4 activation. In this study, TLR4 might be activated by LPS administration. This COX-2 increases PGE2 that leads to induce excitotoxic glutamergic effects and activation of NMDA receptors contribute to depression-like behavior [43]. And, TNF-α also activates NFκB pathway said above and MAPK pathway. MAPK activation leads to phosphorylation of CREB gene in RNA which results in decreased expression of BDNF [44]. Further, anti-inflammatory and analgesic effects of p-CA are quite remarkable in several diseases such as diabetes, gout and arthritis, while p-CA has various bioactivities [45,46]. Its anti-inflammatory and analgesic effects were not inferior to drug such as indomethacin [45]. In this study, anti-depressant-like effect of p-CA is related with the down-regulation of inflammatory cytokines and involved pathways.
Neuroplastic changes are involved in many psychiatric disorders and the key factor in neuroplasticity, BDNF, is associated with clinical symptoms of depression and antidepressant effects [47]. BDNF levels are lower in depressed patients than in healthy subjects, and circulating BDNF significantly increases after antidepressant treatment [48]. In the present study, p-CA improved BDNF levels which had been reduced by LPS. These results suggest that p-CA has antidepressant efficacy by stimulating neuroplasticity as well as minimizing the influence of inflammation.
The clinical relevance of LTD in relation to depression has not been fully revealed yet, but there are a few findings that indicate that LTD is related with depression. LTD is a well-known functional inverse of long-term potentiation (LTP) and a candidate mechanism for learning and memory in the hippocampus [49]. Hippocampal mGlu5 receptors have been shown to mediate depressive-like phenotypes in congenitally learned helpless rats [50]. Ovariectomized rats have demonstrated LTD, the pathological significance of which is not well known, but depression is expressed at higher rates when circulating estrogen levels abruptly change in perimenopausal or postpartum periods [51]. Moreover, it has been reported that inflammation can increase the risk of depression and disrupt LTD [52,53,54,55]. LTD is NMDA receptor-dependent and this requires the opening of NMDA receptors, which leads to Ca2+ influx and subsequent activation of the serine/threonine phosphatase calcineurin/PP2B and PP1. These protein phosphatase cause activation of Bax. Bax was reported to be increased by chronic stress and contribute to depressive behavior [56]. In this study, LPS induced depression-like behavioral changes and disrupts LTD, this suggests a potential association between LTD and depression.
This study demonstrated that LPS potentiates hippocampal LTD, an effect which was prevented by p-CA. Similarly, curcumin on organotypic hippocampal slice cultures attenuates LTD, which is associated with reduced immobility in the FST, upregulation of BDNF, and improvement in deleterious inflammatory factors [4]. Activation of inflammatory cytokines increases or decreases monoaminergic neurotransmitters activation, which results in reuptake of serotonin by presynaptic terminal. For example, previous studies showed that COX-2 mediates the pathway that increases excitatory synaptic transmission [57] and IL-6 decreased inhibitory synaptic transmission such as GABAergic synaptic currents [58]. These effects of cytokines on synaptic transmission are reflected as increased excitability of neurons while p-CA or curcumin reversed this effect.
In this study, the antidepressant and anti-inflammatory effects of p-CA in LPS-treated rats were investigated behaviorally, biologically and electrophysiologically. p-CA improved behavioral despair and motivational behavior, reduced inflammation-related molecular changes, improved neurotrophic activity and reduced hippocampal LTD, which is probably a physiological factor associated with depression. These results indicate that p-CA could potentially alleviate depressive symptoms, including biological aspects as well as behavioral dysfunction such as anhedonia and helplessness. Since p-CA can improve inflammation-related depression-like phenotypes, it may have further benefits for treatment-resistant depression patients.
Figures
{kind=link}

{kind=link}

{kind=link}
{kind=link}

{kind=link}
References
- Sartorius N. The economic and social burden of depression. J Clin Psychiatry 2001;62:8-11.
- Braunstein D, Hardy A, Boucherie Q, Frauger E, Blin O, Gentile G, Micallef J. Antidepressant adherence patterns in older patients: use of a clustering method on a prescription database. Fundam Clin Pharmacol 2017;31:226-236.

- Kirkedal C, Wegener G, Moreira F, Joca SR, Liebenberg N. A dual inhibitor of FAAH and TRPV1 channels shows dose-dependent effect on depression-like behaviour in rats. Acta Neuropsychiatr 2017;29:324-329.
- Choi GY, Kim HB, Hwang ES, Lee S, Kim MJ, Choi JY, Lee SO, Kim SS, Park JH. Curcumin alters neural plasticity and viability of intact hippocampal circuits and attenuates behavioral despair and COX-2 expression in chronically stressed rats. Mediators Inflamm 2017;2017:6280925.
- Su KP. Nutrition, psychoneuroimmunology and depression: the therapeutic implications of omega-3 fatty acids in interferon-α-induced depression. Biomedicine (Taipei) 2015;5:21.
- Reid AM, McNamara JP, Murphy TK, Guzick AG, Storch EA, Goodman WK, Geffken GR, Bussing R. Side-effects of SSRIs disrupt multimodal treatment for pediatric OCD in a randomized-controlled trial. J Psychiatr Res 2015;71:140-147.
- Alcocer-Gómez E, Casas-Barquero N, Williams MR, Romero-Guillena SL, Cañadas-Lozano D, Bullón P, Sánchez-Alcazar JA, Navarro-Pando JM, Cordero MD. Antidepressants induce autophagy dependent-NLRP3-inflammasome inhibition in major depressive disorder. Pharmacol Res 2017;121:114-121.
- Talarowska M, Szemraj J, Gałecki P. The role of interleukin genes in the course of depression. Open Med (Wars) 2016;11:41-48.
- Lavebratt C, Herring MP, Liu JJ, Wei YB, Bossoli D, Hallgren M, Forsell Y. Interleukin-6 and depressive symptom severity in response to physical exercise. Psychiatry Res 2017;252:270-276.
- Wang D, Lin W, Pan Y, Kuang X, Qi X, Sun H. Chronic blockade of glucocorticoid receptors by RU486 enhances lipopolysaccharide-induced depressive-like behaviour and cytokine production in rats. Brain Behav Immun 2011;25:706-714.
- Goshen I, Kreisel T, Ben-Menachem-Zidon O, Licht T, Weidenfeld J, Ben-Hur T, Yirmiya R. Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. Mol Psychiatry 2008;13:717-728.
- Armario A, Hernández J, Bluethmann H, Hidalgo J. IL-6 deficiency leads to increased emotionality in mice: evidence in transgenic mice carrying a null mutation for IL-6. J Neuroimmunol 1998;92:160-169.
- Kaster MP, Gadotti VM, Calixto JB, Santos AR, Rodrigues AL. Depressive-like behavior induced by tumor necrosis factor-α in mice. Neuropharmacology 2012;62:419-426.
- Ming Z, Sawicki G, Bekar LK. Acute systemic LPS-mediated inflammation induces lasting changes in mouse cortical neuromodulation and behavior. Neurosci Lett 2015;590:96-100.
- Li ZQ, Yan ZY, Lan FJ, Dong YQ, Xiong Y. Suppression of NLRP3 inflammasome attenuates stress-induced depression-like behavior in NLGN3-deficient mice. Biochem Biophys Res Commun 2018;501:933-940.
- Wang ML, Yu XJ, Li XG, Pang DZ, Su Q, Saahene RO, Li HB, Mao XY, Liu KL, Fu LY, Li Y, Zhu GQ, Kang YM. Blockade of TLR4 within the paraventricular nucleus attenuates blood pressure by regulating ROS and inflammatory cytokines in prehypertensive rats. Am J Hypertens 2018
- Zhou X, Liu L, Zhang Y, Pu J, Yang L, Zhou C, Yuan S, Zhang H, Xie P. Metabolomics identifies perturbations in amino acid metabolism in the prefrontal cortex of the learned helplessness rat model of depression. Neuroscience 2017;343:1-9.
- Carl H, Walsh E, Eisenlohr-Moul T, Minkel J, Crowther A, Moore T, Gibbs D, Petty C, Bizzell J, Dichter GS, Smoski MJ. Sustained anterior cingulate cortex activation during reward processing predicts response to psychotherapy in major depressive disorder. J Affect Disord 2016;203:204-212.
- Von Frijtag JC, Kamal A, Reijmers LG, Schrama LH, van den Bos R, Spruijt BM. Chronic imipramine treatment partially reverses the long-term changes of hippocampal synaptic plasticity in socially stressed rats. Neurosci Lett 2001;309:153-156.
- Calero P, Jensen SI, Nielsen AT. Broad-host-range ProUSER vectors enable fast characterization of inducible promoters and optimization of p-coumaric acid production in
Pseudomonas putida KT2440. ACS Synth Biol 2016;5:741-753.
- Navaneethan D, Rasool MK. An experimental study to investigate the impact of p-coumaric acid, a common dietary polyphenol, on cadmium chloride-induced renal toxicity. Food Funct 2014;5:2438-2445.
- Kim HB, Lee S, Hwang ES, Maeng S, Park JH. p-Coumaric acid enhances long-term potentiation and recovers scopolamine-induced learning and memory impairments. Biochem Biophys Res Commun 2017;492:493-499.
- Abdel-Moneim A, Yousef AI, Abd El-Twab SM, Abdel Reheim ES, Ashour MB. Gallic acid and p-coumaric acid attenuate type 2 diabetes-induced neurodegeneration in rats. Metab Brain Dis 2017;32:1279-1286.
- Guven M, Aras AB, Akman T, Sen HM, Ozkan A, Salis O, Sehitoglu I, Kalkan Y, Silan C, Deniz M, Cosar M. Neuroprotective effect of p-coumaric acid in rat model of embolic cerebral ischemia. Iran J Basic Med Sci 2015;18:356-363.
- Prasanna N, Rasool M. Modulation of gene-expression profiles associated with sodium arsenite-induced cardiotoxicity by p-coumaric acid, a common dietary polyphenol. J Biochem Mol Toxicol 2014;28:174-180.
- Detke MJ, Rickels M, Lucki I. Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology (Berl) 1995;121:66-72.
- Motaghinejad M, Fatima S, Banifazl S, Bangash MY, Karimian M. Study of the effects of controlled morphine administration for treatment of anxiety, depression and cognition impairment in morphine-addicted rats. Adv Biomed Res 2016;5:178.
- Sadeghi M, Peeri M, Hosseini MJ. Adolescent voluntary exercise attenuated hippocampal innate immunity responses and depressive-like behaviors following maternal separation stress in male rats. Physiol Behav 2016;163:177-183.
- Hwang SH, Lee BH, Kim HJ, Cho HJ, Shin HC, Im KS, Choi SH, Shin TJ, Lee SM, Nam SW, Kim HC, Rhim H, Nah SY. Suppression of metastasis of intravenously-inoculated B16/F10 melanoma cells by the novel ginseng-derived ingredient, gintonin: involvement of autotaxin inhibition. Int J Oncol 2013;42:317-326.
- Zunszain PA, Anacker C, Cattaneo A, Carvalho LA, Pariante CM. Glucocorticoids, cytokines and brain abnormalities in depression. Prog Neuropsychopharmacol Biol Psychiatry 2011;35:722-729.
- Kim JY, Yang SH, Kwon J, Lee HW, Kim H. Mice subjected to uncontrollable electric shocks show depression-like behaviors irrespective of their state of helplessness. Behav Brain Res 2017;322:138-144.
- Yi LT, Liu BB, Li J, Luo L, Liu Q, Geng D, Tang Y, Xia Y, Wu D. BDNF signaling is necessary for the antidepressant-like effect of naringenin. Prog Neuropsychopharmacol Biol Psychiatry 2014;48:135-141.
- Li YC, Liu YM, Shen JD, Chen JJ, Pei YY, Fang XY. Resveratrol ameliorates the depressive-like behaviors and metabolic abnormalities induced by chronic corticosterone injection. Molecules 2016;21:E1341.
- Yi LT, Li CF, Zhan X, Cui CC, Xiao F, Zhou LP, Xie Y. Involvement of monoaminergic system in the antidepressant-like effect of the flavonoid naringenin in mice. Prog Neuropsychopharmacol Biol Psychiatry 2010;34:1223-1228.
- Ali SH, Madhana RM, K V A, Kasala ER, Bodduluru LN, Pitta S, Mahareddy JR, Lahkar M. Resveratrol ameliorates depressive-like behavior in repeated corticosterone-induced depression in mice. Steroids 2015;101:37-42.
- Yalcin I, Belzung C, Surget A. Mouse strain differences in the unpredictable chronic mild stress: a four-antidepressant survey. Behav Brain Res 2008;193:140-143.
- Brachman RA, McGowan JC, Perusini JN, Lim SC, Pham TH, Faye C, Gardier AM, Mendez-David I, David DJ, Hen R, Denny CA. Ketamine as a prophylactic against stress-induced depressive-like behavior. Biol Psychiatry 2016;79:776-786.
- Miller AH, Maletic V, Raison CL. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry 2009;65:732-741.
- Dowlati Y, Herrmann N, Swardfager W, Liu H, Sham L, Reim EK, Lanctôt KL. A meta-analysis of cytokines in major depression. Biol Psychiatry 2010;67:446-457.
- Müller N, Schwarz MJ, Dehning S, Douhe A, Cerovecki A, Goldstein-Müller B, Spellmann I, Hetzel G, Maino K, Kleindienst N, Möller HJ, Arolt V, Riedel M. The cyclooxygenase-2 inhibitor celecoxib has therapeutic effects in major depression: results of a double-blind, randomized, placebo controlled, add-on pilot study to reboxetine. Mol Psychiatry 2006;11:680-684.
- Myint AM, Steinbusch HW, Goeghegan L, Luchtman D, Kim YK, Leonard BE. Effect of the COX-2 inhibitor celecoxib on behavioural and immune changes in an olfactory bulbectomised rat model of depression. Neuroimmunomodulation 2007;14:65-71.
- Maes M, Bosmans E, De Jongh R, Kenis G, Vandoolaeghe E, Neels H. Increased serum IL-6 and IL-1 receptor antagonist concentrations in major depression and treatment resistant depression. Cytokine 1997;9:853-858.
- Kubera M, Obuchowicz E, Goehler L, Brzeszcz J, Maes M. In animal models, psychosocial stress-induced (neuro) inflammation, apoptosis and reduced neurogenesis are associated to the onset of depression. Prog Neuropsychopharmacol Biol Psychiatry 2011;35:744-759.
- Gass P, Riva MA. CREB, neurogenesis and depression. BioEssays 2007;29:957-961.
- Pei K, Ou J, Huang J, Ou S. p-Coumaric acid and its conjugates: dietary sources, pharmacokinetic properties and biological activities. J Sci Food Agric 2016;96:2952-2962.
- Zhu H, Liang QH, Xiong XG, Wang Y, Zhang ZH, Sun MJ, Lu X, Wu D. Anti-inflammatory effects of p-coumaric acid, a natural compound of
Oldenlandia diffusa , on arthritis model rats. Evid Based Complement Alternat Med 2018;2018:5198594.
- Brunoni AR, Lopes M, Fregni F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: implications for the role of neuroplasticity in depression. Int J Neuropsychopharmacol 2008;11:1169-1180.
- Aydemir C, Yalcin ES, Aksaray S, Kisa C, Yildirim SG, Uzbay T, Goka E. Brain-derived neurotrophic factor (BDNF) changes in the serum of depressed women. Prog Neuropsychopharmacol Biol Psychiatry 2006;30:1256-1260.
- Bear MF, Malenka RC. Synaptic plasticity: LTP and LTD. Curr Opin Neurobiol 1994;4:389-399.
- Pignatelli M, Vollmayr B, Richter SH, Middei S, Matrisciano F, Molinaro G, Nasca C, Battaglia G, Ammassari-Teule M, Feligioni M, Nisticò R, Nicoletti F, Gass P. Enhanced mGlu5-receptor dependent long-term depression at the Schaffer collateral-CA1 synapse of congenitally learned helpless rats. Neuropharmacology 2013;66:339-347.
- Kim HB, Kwon BJ, Cho HJ, Kim JW, Chon JW, Do MH, Park SY, Kim SY, Maeng SH, Park YK, Park JH. Long-term treatment with oriental medicinal herb artemisia princeps alters neuroplasticity in a rat model of ovarian hormone deficiency. Exp Neurobiol 2015;24:71-83.
- Korczak DJ, Pereira S, Koulajian K, Matejcek A, Giacca A. Type 1 diabetes mellitus and major depressive disorder: evidence for a biological link. Diabetologia 2011;54:2483-2493.
- Dickens C, McGowan L, Clark-Carter D, Creed F. Depression in rheumatoid arthritis: a systematic review of the literature with meta-analysis. Psychosom Med 2002;64:52-60.
- Benros ME, Waltoft BL, Nordentoft M, Ostergaard SD, Eaton WW, Krogh J, Mortensen PB. Autoimmune diseases and severe infections as risk factors for mood disorders: a nationwide study. JAMA Psychiatry 2013;70:812-820.
- Min SS, Quan HY, Ma J, Lee KH, Back SK, Na HS, Han SH, Yee JY, Kim C, Han JS, Seol GH. Impairment of long-term depression induced by chronic brain inflammation in rats. Biochem Biophys Res Commun 2009;383:93-97.
- Liu X, Gu QH, Duan K, Li Z. NMDA receptor-dependent LTD is required for consolidation but not acquisition of fear memory. J Neurosci 2014;34:8741-8748.
- Li B, Guo L, Ku T, Chen M, Li G, Sang N. PM2.5 exposure stimulates COX-2-mediated excitatory synaptic transmission via ROS-NF-κB pathway. Chemosphere 2018;190:124-134.
- Cuevas-Olguin R, Esquivel-Rendon E, Vargas-Mireles J, Garcia-Oscos F, Miranda-Morales M, Salgado H, Rose-John S, Atzori M. Interleukin 6 trans-signaling regulates basal synaptic transmission and sensitivity to pentylenetetrazole-induced seizures in mice. Synapse 2017;71:e21984.