Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2020; 29(2): 138-149
Published online April 30, 2020
https://doi.org/10.5607/en19072
© The Korean Society for Brain and Neural Sciences
Loss of MicroRNA-137 Impairs the Homeostasis of Potassium in Neurons via KCC2
Ting-Wei Mi1,2†, Xiao-Wen Sun1,3†, Zhi-Meng Wang1,3†, Ying-Ying Wang1,3†, Xuan-Cheng He1,2†, Cong Liu1,3, Shuang-Feng Zhang1,4, Hong-Zhen Du1,2, Chang-Mei Liu1,2,3* and Zhao-Qian Teng1,2,3*
1State Key Laboratory of Stem Cell and Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, 2Institute for Stem Cell and Regeneration, Chinese Academy of Sciences, Beijing 100101, 3Savaid Medical School, University of Chinese Academy of Sciences, Beijing 100049, 4School of Life Sciences, University of Science and Technology of China, Hefei 230026, China
Correspondence to: *To whom correspondence should be addressed.
Zhao-Qian Teng, TEL: 86-10-82619699, FAX: 86-10-64807099
e-mail: tengzq@ioz.ac.cn
Chang-Mei Liu, TEL: 86-10-82619690, FAX: 86-10-64807316
e-mail: liuchm@ioz.ac.cn
†These authors equally contributed to this work.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License
(http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and
reproduction in any medium, provided the original work is properly cited.
Abstract
Neuropsychiatric disorders are the leading cause of mental and intellectual disabilities worldwide. Current therapies against neuropsychiatric disorders are very limited, and very little is known about the onset and development of these diseases, and their most effective treatments.
Graphical Abstract
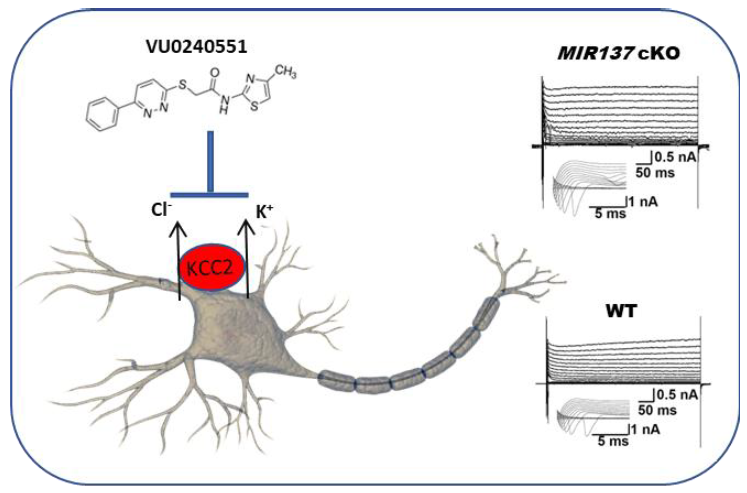
Keywords:
INTRODUCTION
Neuropsychiatric disorders, including schizophrenia, autism, depression, and anxiety, are very common all around the world [1]. The limited effectiveness of current therapies against neuropsychiatric disorders and neurological disorders highlights the urgent need for understanding their pathological mechanisms and for developing new approaches to prevent or retard the disease progression [2].
Genome-wide association studies (GWAS) have identified
Neural activity depends on electric signals that are transmitted from the presynaptic neuron to the postsynaptic cell via chemical signaling. The positive or negative change in membrane potential of the postsynaptic neuron is caused by the activation of postsynaptic receptors, which are ion channels whose activation alters permeability for specific ions [13]. Although genetic variation in genes coding for ion channels increases risk for psychiatric disorders [14-17], little is known about the function of miR-137 on ion channels in neurons.
In this study, we provide the first evidence that loss of miR-137 results in impaired homeostasis of potassium in neurons, both
MATERIALS AND METHODS
Mice
All experiments involving animals were performed in accordance with the animal protocol approved by the Institutional Animal Care and Use Committee at the Institute of Zoology, Chinese Academy of Sciences. Mice were housed in groups of 3~5 animals under a 12 h light/12 h dark cycle, and were fed ad libitum on a standard mouse diet. The miR-137f/f mice were generated as previously described [10]. The Emx1-Cre transgenic mice were bought from Jackson Laboratory (Stock No. 005628). The miR-137 conditional knockout mice were generated by breeding miR-137f/f mice with Emx1-Cre transgenic mice, as described previously [19].
Primary hippocampal neuron culture
Hippocampal neurons were isolated from P0
Dual luciferase assays
Approximately 300 base pairs around the predicted target site from the KCC2 3’UTR was cloned into the pIS2 vector using the XhoI and NotI restriction sites in the multiple cloning region downstream of the luciferase reporter gene. Mutagenesis of the binding site on KCC2 3’UTR was performed using the QuickChange II Site-directed Mutagenesis Kit (Stratagene, La Jolla, CA) according to the manufacturer's protocol. All plasmid clones were then verified by sequencing.
Dual luciferase transfection assays were performed as previously described [20, 21]. In brief, HEK293 cells in 24-well plates were transfected with sh-miR-137 (pCR2.1 TOPO vector) and pIS2-3’UTR or mutated pIS2-3’UTR using Lipofectamine 2000 (Invitrogen). Meanwhile, pIS2 vector with no 3’UTR was cotranfected with U6-neg-shRNA (pCR2.1 TOPO vector) or sh-miR-137 to set up as a control. All Luciferase readings were recorded using Dual-Luciferase Reporter 1000 System (Promega) following manufacturer's instructions.
Electrophysiological recordings
Whole-cell patch-clamp recordings were carried out using an Axopatch 700B amplifier (Axon Instruments, Union City, CA). The pClamp10.6 software was used for data acquisition and analysis. Patch pipettes (6~10 MΩ) were pulled from borosilicate glass capillaries with a micropipette puller (Sutter instrument, USA). The internal pipette solution contained (in mM): 135 K-gluconate, 10 HEPES, 2 MgCl2, 10 EGTA, 0.3 MgGTP, and 0.5 Na2ATP (pH 7.3 with KOH). The membrane potential was held at -65 mV. Series resistances and cell capacitance compensation were carried out prior to recording. The recordings were included only in those with high resistance seal (>1 GΩ) and a series resistance <25 MΩ.
RNA extraction and qRT-PCR
Total RNA was extracted from hippocampus tissue or cultured neurons using TRIzol reagent (Invitrogen). Two micrograms of total RNA were reverse transcribed with either oligo (dT) primers or specific primers by a Transcriptor First Strand cDNA Synthesis Kit (Roche). For qRT-PCR analysis, 25 ng of cDNA and 0.5 µM primers were used in a final volume of 20 µl according to the manufacturer’s instructions (SYBR Green Master, Roche). Each reaction was run in triplicate and analyzed following the △△CT method using U6 or GAPDH as a normalization control. The following primers are used: KCC2 (forward: 5’-GGGCAGAGAGTACGATGGC-3’; reverse: 5’-TGGGGTAGGTTGGTGTAGTTG-3’), GAPDH (forward: 5’-AAGGTCATCCCAGAGCTGAA-3’; reverse: 5’-AGGAGACAACCTGGTCCTCA-3’).
Protein quantification
Hippocampal tissues or cultured neurons were lysed in a buffer containing 25 mM HEPES at pH7.9, 150 mM NaCl, 1 mM PMSF, 20 mM NaF, 1 mM DTT, 0.1% NP40, and proteinase inhibitor cocktails (Roche). Protein concentrations were determined by Folin phenol method with bovine serum albumin as protein standard. Twenty micrograms of protein were separated on 8~12% SDS-PAGE gels (Bio-Rad) and transferred to PVDF membranes (Millipore). The membranes were then blocked in 5% BSA in TBS-T with 0.05% Tween-20, and incubated with primary antibodies at 4°C overnights. Dilutions of primary antibodies were 1:1,000 for KCC2 (Millipore, #07–432), and 1:10,000 for β-actin antibody (Sigma). As for the secondary antibodies, we used HRP-linked goat anti-rabbit at 1:500. Enhanced chemo luminescence (ECL, Pierce) was used for detection. Quantifications of Western blots were determined using Quantity One V4.4.0 (BioRad).
Immunohistochemistry
Mice were anesthetized and transcardially perfused with cold PBS, followed by 4% PFA in PBS (pH 7.4). Brain tissue was dissected out, equilibrated in 30% sucrose, and sectioned into 40 µm-thick sections. The brain sections were washed in PBS for 15 min three times, and then blocked in a blocking solution (3% BSA, 0.3%Triton X-100, 0.2% sodium azide) at room temperature for 1 h. The primary antibodies we used were as follows: anti-KCC2 (1:1,000, Millipore, #07–432), anti-Map2 (1:1,000, Millipore, Mab3418). After overnight incubation with primary antibody at 4°C, the brain sections were washed with TBS for 30 min three times and then incubated with the secondary antibodies conjugated with Alexa Fluor 488 or 594 (1:500). Sections were finally stained with DAPI and mounted on glass-slides using adhesion anti-fade medium.
Lentivirus production and in vivo grafting
KCC2 shRNA sequence (CUACGAGAA GACAUUAGUA) [22] was inserted in the U6-shRNA lentiviral construct. Lenti-sh-KCC2 and lenti-sh-Neg (negative control) viruses were produced with titers at a range around 1×109 TU/ml as described previously [23, 24]. Lentivirus was grafted stereotaxically into the hippocampus of 8-week-old male
Behavioral tests
Mice were kept in groups of 4~5 animals on a 12:12 h light:dark cycle. The open field test and the light-dark preference test were performed during the light phase at week 3 after lentiviral injection as previously described [19]. Videos were recorded and analyzed by the software Smart V3.0.03 (Panlab, Barcelona, Spain).
Statistical analyses
Either unpaired Student’s two-tailed t tests or ANOVA with Tukey’s post hoc tests were conducted using IBM SPSS Statistics V26 software. Samples sizes were provided in each figure legend. All data were presented as mean±SEM. Differences were considered statistically significant when p<0.05.
RESULTS
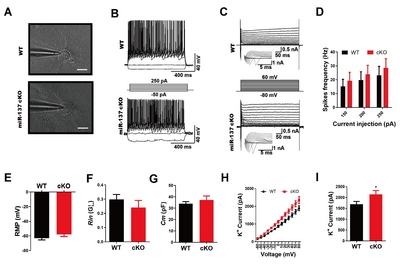
Loss of miR-137 impairs the homeostasis of potassium in primary hippocampal neurons
We originally generated miR-137 conditional knockout mice that displayed dysregulated synaptic plasticity, repetitive behavior, anxiety-like behavior, and impaired learning and social behavior [10]. Since neurological and neuropsychological diseases are pathophysiologically linked to potassium channel dysfunction [26, 27], we speculated that loss of miR-137 may play a role in regulating K+ currents and thus result in neurodysfunction.
To examine the electrophysiological properties of primary hippocampal neurons isolated from
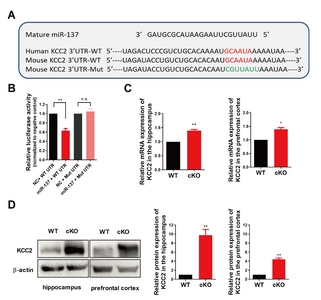
K+–Cl- cotransporter 2 (KCC2) is a direct downstream target of miR-137
To get more insight into the molecular mechanism underlying the effect of miR-137 loss-of-function on voltage-gated K+ current, we performed a combined computational and experimental study to identify the downstream targets of miR-137 in the brain. We first used the TargetScan program to predict mRNA targets [28]. TargetScan analysis identified 15 potassium channel associated targets predicted to be responsive to miR-137 and conserved among species. Among these candidate targets, the K+–Cl- cotransporter 2 (aka: KCC2 and SLC12A5) is known to play pivotal roles in the physiology of neurons, and its malfunction has been linked to multiple neurological diseases including seizures, epilepsy, and schizophrenia [29-33]. Indeed, there is a highly conserved binding site of miR-137 on the 3’-UTR sequence of mouse
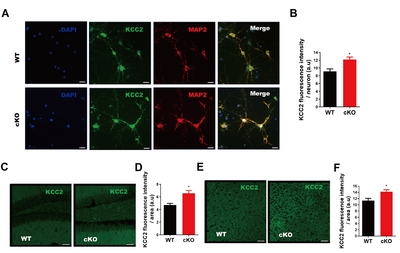
Next, we examined the expression levels of KCC2 in
KCC2 antagonist maintains potassium homeostasis in MIR137 cKO neurons
To further explore the role of KCC2 in the electrophysiological properties of neurons, we used a pharmacological agent, the KCC2 antagonist VU0240551, to intervene the function of KCC2 by preincubating neurons with the agent for 1 hour (10 µM), and performed the whole-cell patch-clamp recordings of primary hippocampal neurons at DIV 14. All primary hippocampal neurons elicited multiple action potentials upon the injection of depolarizing currents (Fig. 4A) and large fast-inactivating inward currents followed by outward potassium currents when evoked by a series of voltage steps (Fig. 4B). We then assessed membrane properties of hippocampal neurons that threated with VU0240551 by recording voltage-dependent currents in voltage-clamp mode. We found that maximum peak outward potassium amplitude of VU0240551-treated
Knockdown of KCC2 ameliorates anxiety-like behavior in MIR137 cKO mice
Recently, we found that mice with forebrain-specific miR-137 loss-of-function can survive to adulthood, but exhibit anxiety-like behavior [19]. To examine whether knockdown of KCC2 might be beneficial to ameliorate anxiety-like behavior in
In the open field test, sh-KCC2 significantly ameliorates the anxiety-like behavior in
DISCUSSION
Dysfunction of miR-137 has been linked with the pathogenesis of schizophrenia [3], bipolar disorder [4], anxiety and depression [19], and autism spectrum disorders [5, 10]. Fine-tuning the expression of miR-137 is critical in regulating neural development and synaptic plasticity [9, 20, 34]. Overexpression of miR-137 results in changes in synaptic vesicle pool distribution, impaired induction of mossy fiber long-term potentiation and deficits in hippocampus-dependent learning and memory [9]. He et al. [34] then confirmed these observed changes in synaptic transmission upon miR-137 overexpression. Although selective synaptic vesicle docking defects were not obtained, miR-137 overexpression had remarkable effects on docking, active zone length and total vesicle number [34]. Syt1, complexin-1 and neuroligin-3 are known miR-137 targets involved in synapse development [9, 34]. In contrast, a complete knockout of
KCC2 mutations or dysfunctions have been identified as a critical component in the development of autism spectrum disorder [37], schizophrenia, epilepsy and seizures [38-42], neuropathic pain [43]. KCC2 has been well-known for its role in maintaining a low intracellular Cl- concentration ([Cl-]i) essential for hyperpolarizing inhibition mediated by GABAA receptors [44], its loss-of-function results in enhanced [Cl-]i which thereby represses the inhibitory strength of GABA and glycine, whose cognate receptors are ligand-gated ion channels permeable to Cl- and HCO3- ions [45-47]. Besides its central role in hyperpolarizing inhibitory signaling based on chloride currents which are mediated by GABA- or glycine-gated receptor channels, KCC2 also acts a structural protein crucially involved in the maturation and regulation of excitatory glutamatergic synapses [48-51]. KCC2 is required for neuronal maturation by rendering GABA hyperpolarizing [49,52-54], and overexpression of KCC2 enhances dendritic spines in the adult nervous system in mice [55]. Therefore, we speculate that the elevated expression of KCC2 may also contribute to dysregulated synaptic plasticity and altered behaviors in
Potassium is essential for the proper function of all cells [56]. In neurons, the sodium-potassium flux generates the electrical potential that aids the conduction of nerve impulses [56]. Potassium channels participate in cell ionic balance and serve the fundamental function of supporting action potentials and electrical signal propagation along the neurons and their myelinated axons [57-59]. The increased outward K+ currents in
Surprisingly, increased K+ currents in
In summary, we show here that miR-137 is a crucial player in the homeostasis of potassium by directly targeting KCC2, and treatment with the KCC2 antagonist can maintain potassium homeostasis in
ACKNOWLEDGEMENTS
This work was supported by grants from the National Key Research and Development Program of China Project (2018YFA0108001), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA16010300), the National Science Foundation of China (91753140), the Beijing Natural Science Foundation (7182107), and the Open Project Program of State Key Laboratory of Stem Cell and Reproductive Biology.
Figures
{kind=link}
MiR-137 deficiency impairs K+ efflux of potassium in primary mouse hippocampal neurons. (A) Phase-contrast images of primary mouse hippocampal neurons during patch clamp recordings at DIV 14. (B) Representative traces of action potentials in response to step current injections in primary mouse hippocampal neurons at DIV 14. Membrane potential was maintained at approximately -40 mV. Step currents were injected from -50 pA to +250 pA in 50 pA increments (middle panel). All neurons elicited multiple action potentials upon the injection of depolarizing currents. (C) Representative traces of whole-cell currents in voltage-clamp mode. Primary neurons from
{kind=link}
KCC2 is a direct target of miR-137. (A) Sequence alignment of miR-137 and the KCC2 3’-UTR, which contains a predicted conserved miR-137-biding site. The seed-recognizing site in the KCC2 3’-UTR is indicated in red, while the mutant KCC2 3’-UTR site is denoted in green. (B) KCC2 WT 3’-UTR-dependent expression of a Renilla luciferase was reduced, while mutation of the miR-137 binding site in the KCC2 3’-UTR did not affect the Renilla luciferase activity. (C) KCC2 mRNA expression levels were upregulated both in the hippocampus and in the prefrontal cortex of
{kind=link}
KCC2 is upregulated upon the loss of miR-137 both
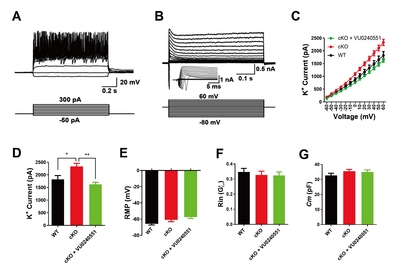
{kind=link}
KCC2 antagonist maintains potassium homeostasis in mouse
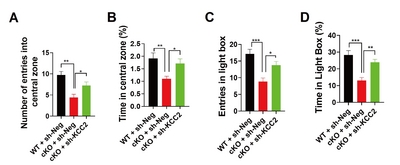
{kind=link}
KCC2 knockdown rescues anxiety-like behaviors in
References
- Barešić A, Nash AJ, Dahoun T, Howes O, Lenhard B (2020) Understanding the genetics of neuropsychiatric disorders: the potential role of genomic regulatory blocks. Mol Psychiatry 25:6-18.



- Lee HM, Kim Y (2016) Drug repurposing is a new opportunity for developing drugs against neuropsychiatric disorders. Schizophr Res Treatment 2016:6378137.
- Ripke S, O'Dushlaine C, Chambert K, Moran JL, Kähler AK, Akterin S, Bergen SE, Collins AL, Crowley JJ, Fromer M, Kim Y, Lee SH, Magnusson PK, Sanchez N, Stahl EA, Williams S, Wray NR, Xia K, Bettella F, Borglum AD, Bulik-Sullivan BK, Cormican P, Craddock N, de Leeuw C, Durmishi N, Gill M, Golimbet V, Hamshere ML, Holmans P, Hougaard DM, Kendler KS, Lin K, Morris DW, Mors O, Mortensen PB, Neale BM, O'Neill FA, Owen MJ, Milovancevic MP, Posthuma D, Powell J, Richards AL, Riley BP, Ruderfer D, Rujescu D, Sigurdsson E, Silagadze T, Smit AB, Stefansson H, Steinberg S, Suvisaari J, Tosato S, Verhage M, Walters JT; Multicenter Genetic Studies of Schizophrenia Consortium, Levinson DF, Gejman PV, Kendler KS, Laurent C, Mowry BJ, O'Donovan MC, Owen MJ, Pulver AE, Riley BP, Schwab SG, Wildenauer DB, Dudbridge F, Holmans P, Shi J, Albus M, Alexander M, Campion D, Cohen D, Dikeos D, Duan J, Eichhammer P, Godard S, Hansen M, Lerer FB, Liang KY, Maier W, Mallet J, Nertney DA, Nestadt G, Norton N, O'Neill FA, Papadimitriou GN, Ribble R, Sanders AR, Silverman JM, Walsh D, Williams NM, Wormley B; Psychosis Endophenotypes International Consortium, Arranz MJ, Bakker S, Bender S, Bramon E, Collier D, Crespo-Facorro B, Hall J, Iyegbe C, Jablensky A, Kahn RS, Kalaydjieva L, Lawrie S, Lewis CM, Lin K, Linszen DH, Mata I, McIntosh A, Murray RM, Ophoff RA, Powell J, Rujescu D, Van Os J, Walshe M, Weisbrod M, Wiersma D; Wellcome Trust Case Control Consortium 2, Donnelly P, Barroso I, Blackwell JM, Bramon E, Brown MA, Casas JP, Corvin AP, Deloukas P, Duncanson A, Jankowski J, Markus HS, Mathew CG, Palmer CN, Plomin R, Rautanen A, Sawcer SJ, Trembath RC, Viswanathan AC, Wood NW, Spencer CC, Band G, Bellenguez C, Freeman C, Hellenthal G, Giannoulatou E, Pirinen M, Pearson RD, Strange A, Su Z, Vukcevic D, Donnelly P, Langford C, Hunt SE, Edkins S, Gwilliam R, Blackburn H, Bumpstead SJ, Dronov S, Gillman M, Gray E, Hammond N, Jayakumar A, McCann OT, Liddle J, Potter SC, Ravindrarajah R, Ricketts M, Tashakkori-Ghanbaria A, Waller MJ, Weston P, Widaa S, Whittaker P, Barroso I, Deloukas P, Mathew CG, Blackwell JM, Brown MA, Corvin AP, McCarthy MI, Spencer CC, Bramon E, Corvin AP, O'Donovan MC, Stefansson K, Scolnick E, Purcell S, McCarroll SA, Sklar P, Hultman CM, Sullivan PF (2013) Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat Genet 45:1150-1159.
- Duan J, Shi J, Fiorentino A, Leites C, Chen X, Moy W, Chen J, Alexandrov BS, Usheva A, He D, Freda J, O'Brien NL; Molecular Genetics of Schizophrenia collaboration; Genomic Psychiatric Cohort consortium, McQuillin A, Sanders AR, Gershon ES, DeLisi LE, Bishop AR, Gurling HM, Pato MT, Levinson DF, Kendler KS, Pato CN, Gejman PV (2014) A rare functional noncoding variant at the GWAS-implicated MIR137/MIR2682 locus might confer risk to schizophrenia and bipolar disorder. Am J Hum Genet 95:744-753.
- Pinto D, Delaby E, Merico D, Barbosa M, Merikangas A, Klei L, Thiruvahindrapuram B, Xu X, Ziman R, Wang Z, Vorstman JA, Thompson A, Regan R, Pilorge M, Pellecchia G, Pagnamenta AT, Oliveira B, Marshall CR, Magalhaes TR, Lowe JK, Howe JL, Griswold AJ, Gilbert J, Duketis E, Dombroski BA, De Jonge MV, Cuccaro M, Crawford EL, Correia CT, Conroy J, Conceição IC, Chiocchetti AG, Casey JP, Cai G, Cabrol C, Bolshakova N, Bacchelli E, Anney R, Gallinger S, Cotterchio M, Casey G, Zwaigenbaum L, Wittemeyer K, Wing K, Wallace S, van Engeland H, Tryfon A, Thomson S, Soorya L, RogéB , Roberts W, Poustka F, Mouga S, Minshew N, McInnes LA, McGrew SG, Lord C, Leboyer M, Le Couteur AS, Kolevzon A, Jiménez González P, Jacob S, Holt R, Guter S, Green J, Green A, Gillberg C, Fernandez BA, Duque F, Delorme R, Dawson G, Chaste P, CaféC , Brennan S, Bourgeron T, Bolton PF, Bölte S, Bernier R, Baird G, Bailey AJ, Anagnostou E, Almeida J, Wijsman EM, Vieland VJ, Vicente AM, Schellenberg GD, Pericak-Vance M, Paterson AD, Parr JR, Oliveira G, Nurnberger JI, Monaco AP, Maestrini E, Klauck SM, Hakonarson H, Haines JL, Geschwind DH, Freitag CM, Folstein SE, Ennis S, Coon H, Battaglia A, Szatmari P, Sutcliffe JS, Hallmayer J, Gill M, Cook EH, Buxbaum JD, Devlin B, Gallagher L, Betancur C, Scherer SW (2014) Convergence of genes and cellular pathways dysregulated in autism spectrum disorders. Am J Hum Genet 94:677-694.
- Agarwal V, Bell GW, Nam JW, Bartel DP (2015) Predicting effective microRNA target sites in mammalian mRNAs. Elife 4:e05005.
- Szulwach KE, Li X, Smrt RD, Li Y, Luo Y, Lin L, Santistevan NJ, Li W, Zhao X, Jin P (2010) Cross talk between microRNA and epigenetic regulation in adult neurogenesis. J Cell Biol 189:127-141.
- Smrt RD, Szulwach KE, Pfeiffer RL, Li X, Guo W, Pathania M, Teng ZQ, Luo Y, Peng J, Bordey A, Jin P, Zhao X (2010) MicroRNA miR-137 regulates neuronal maturation by targeting ubiquitin ligase mind bomb-1. Stem Cells 28:1060-1070.
- Siegert S, Seo J, Kwon EJ, Rudenko A, Cho S, Wang W, Flood Z, Martorell AJ, Ericsson M, Mungenast AE, Tsai LH (2015) The schizophrenia risk gene product miR-137 alters presynaptic plasticity. Nat Neurosci 18:1008-1016.
- Cheng Y, Wang ZM, Tan W, Wang X, Li Y, Bai B, Li Y, Zhang SF, Yan HL, Chen ZL, Liu CM, Mi TW, Xia S, Zhou Z, Liu A, Tang GB, Liu C, Dai ZJ, Wang YY, Wang H, Wang X, Kang Y, Lin L, Chen Z, Xie N, Sun Q, Xie W, Peng J, Chen D, Teng ZQ, Jin P (2018) Partial loss of psychiatric risk gene Mir137 in mice causes repetitive behavior and impairs sociability and learning via increased Pde10a. Nat Neurosci 21:1689-1703.
- Thomas KT, Anderson BR, Shah N, Zimmer SE, Hawkins D, Valdez AN, Gu Q, Bassell GJ (2017) Inhibition of the schizophrenia-associated microRNA miR-137 disrupts Nrg1¥á neurodevelopmental signal transduction. Cell Rep 20:1-12.
- Wright C, Turner JA, Calhoun VD, Perrone-Bizzozero N (2013) Potential impact of miR-137 and its targets in schizophrenia. Front Genet 4:58.
- Schulte JT, Wierenga CJ, Bruining H (2018) Chloride transporters and GABA polarity in developmental, neurological and psychiatric conditions. Neurosci Biobehav Rev 90:260-271.
- Moon AL, Haan N, Wilkinson LS, Thomas KL, Hall J (2018) CACNA1C: association with psychiatric disorders, behavior, and neurogenesis. Schizophr Bull 44:958-965.
- Verkman AS, Smith AJ, Phuan PW, Tradtrantip L, Anderson MO (2017) The aquaporin-4 water channel as a potential drug target in neurological disorders. Expert Opin Ther Targets 21:1161-1170.
- Kabir ZD, Martínez-Rivera A, Rajadhyaksha AM (2017) From gene to behavior: L-type calcium channel mechanisms underlying neuropsychiatric symptoms. Neurotherapeutics 14:588-613.
- Pinggera A, Striessnig J (2016) Cav 1.3 (CACNA1D) L-type Ca2+ channel dysfunction in CNS disorders. J Physiol 594:5839-5849.
- Deisz RA, Wierschke S, Schneider UC, Dehnicke C (2014) Effects of VU0240551, a novel KCC2 antagonist, and DIDS on chloride homeostasis of neocortical neurons from rats and humans. Neuroscience 277:831-841.
- Yan HL, Sun XW, Wang ZM, Liu PP, Mi TW, Liu C, Wang YY, He XC, Du HZ, Liu CM, Teng ZQ (2019) MiR-137 deficiency causes anxiety-like behaviors in mice. Front Mol Neurosci 12:260.
- Chen L, Wang X, Huang W, Ying T, Chen M, Cao J, Wang M (2017) MicroRNA-137 and its downstream target LSD1 inversely regulate anesthetics-induced neurotoxicity in dorsal root ganglion neurons. Brain Res Bull 135:1-7.
- Liu C, Teng ZQ, Santistevan NJ, Szulwach KE, Guo W, Jin P, Zhao X (2010) Epigenetic regulation of miR-184 by MBD1 governs neural stem cell proliferation and differentiation. Cell Stem Cell 6:433-444.
- Ivakine EA, Acton BA, Mahadevan V, Ormond J, Tang M, Pressey JC, Huang MY, Ng D, Delpire E, Salter MW, Woodin MA, McInnes RR (2013) Neto2 is a KCC2 interacting protein required for neuronal Cl- regulation in hippocampal neurons. Proc Natl Acad Sci U S A 110:3561-3566.
- Liu PP, Tang GB, Xu YJ, Zeng YQ, Zhang SF, Du HZ, Teng ZQ, Liu CM (2017) MiR-203 interplays with polycomb repressive complexes to regulate the proliferation of neural stem/progenitor cells. Stem Cell Reports 9:190-202.
- Liu C, Dai SK, Sun Z, Wang Z, Liu PP, Du HZ, Yu S, Liu CM, Teng ZQ (2019) GA-binding protein GABPβ1 is required for the proliferation of neural stem/progenitor cells. Stem Cell Res 39:101501.
- Heldt SA, Stanek L, Chhatwal JP, Ressler KJ (2007) Hippocampus-specific deletion of BDNF in adult mice impairs spatial memory and extinction of aversive memories. Mol Psychiatry 12:656-670.
- Kahle KT, Khanna AR, Alper SL, Adragna NC, Lauf PK, Sun D, Delpire E (2015) K-Cl cotransporters, cell volume homeostasis, and neurological disease. Trends Mol Med 21:513-523.
- Benatar M (2000) Neurological potassium channelopathies. QJM 93:787-797.
- Lewis BP, Burge CB, Bartel DP (2005) Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 120:15-20.
- Chen L, Wan L, Wu Z, Ren W, Huang Y, Qian B, Wang Y (2017) KCC2 downregulation facilitates epileptic seizures. Sci Rep 7:156.
- Di Cristo G, Awad PN, Hamidi S, Avoli M (2018) KCC2, epileptiform synchronization, and epileptic disorders. Prog Neurobiol 162:1-16.
- Hübner CA (2014) The KCl-cotransporter KCC2 linked to epilepsy. EMBO Rep 15:732-733.
- Moore YE, Deeb TZ, Chadchankar H, Brandon NJ, Moss SJ (2018) Potentiating KCC2 activity is sufficient to limit the onset and severity of seizures. Proc Natl Acad Sci U S A 115:10166-10171.
- Moore YE, Kelley MR, Brandon NJ, Deeb TZ, Moss SJ (2017) Seizing control of KCC2: a new therapeutic target for epilepsy. Trends Neurosci 40:555-571.
- He E, Lozano MAG, Stringer S, Watanabe K, Sakamoto K, den Oudsten F, Koopmans F, Giamberardino SN, Hammerschlag A, Cornelisse LN, Li KW, van Weering J, Posthuma D, Smit AB, Sullivan PF, Verhage M (2018) MIR137 schizophrenia-associated locus controls synaptic function by regulating synaptogenesis, synapse maturation and synaptic transmission. Hum Mol Genet 27:1879-1891.
- Crowley JJ, Collins AL, Lee RJ, Nonneman RJ, Farrell MS, Ancalade N, Mugford JW, Agster KL, Nikolova VD, Moy SS, Sullivan PF (2015) Disruption of the microRNA 137 primary transcript results in early embryonic lethality in mice. Biol Psychiatry 77:e5-e7.
- Tillman L, Zhang J (2019) Crossing the chloride channel: the current and potential therapeutic value of the neuronal K+-Cl- cotransporter KCC2. Biomed Res Int 2019:8941046.
- Merner ND, Chandler MR, Bourassa C, Liang B, Khanna AR, Dion P, Rouleau GA, Kahle KT (2015) Regulatory domain or CpG site variation in SLC12A5, encoding the chloride transporter KCC2, in human autism and schizophrenia. Front Cell Neurosci 9:386.
- Puskarjov M, Seja P, Heron SE, Williams TC, Ahmad F, Iona X, Oliver KL, Grinton BE, Vutskits L, Scheffer IE, Petrou S, Blaesse P, Dibbens LM, Berkovic SF, Kaila K (2014) A variant of KCC2 from patients with febrile seizures impairs neuronal Cl- extrusion and dendritic spine formation. EMBO Rep 15:723-729.
- Kahle KT, Merner ND, Friedel P, Silayeva L, Liang B, Khanna A, Shang Y, Lachance-Touchette P, Bourassa C, Levert A, Dion PA, Walcott B, Spiegelman D, Dionne-Laporte A, Hodgkinson A, Awadalla P, Nikbakht H, Majewski J, Cossette P, Deeb TZ, Moss SJ, Medina I, Rouleau GA (2014) Genetically encoded impairment of neuronal KCC2 cotransporter function in human idiopathic generalized epilepsy. EMBO Rep 15:766-774.
- Stödberg T, McTague A, Ruiz AJ, Hirata H, Zhen J, Long P, Farabella I, Meyer E, Kawahara A, Vassallo G, Stivaros SM, Bjursell MK, Stranneheim H, Tigerschiöld S, Persson B, Bangash I, Das K, Hughes D, Lesko N, Lundeberg J, Scott RC, Poduri A, Scheffer IE, Smith H, Gissen P, Schorge S, Reith ME, Topf M, Kullmann DM, Harvey RJ, Wedell A, Kurian MA (2015) Mutations in SLC12A5 in epilepsy of infancy with migrating focal seizures. Nat Commun 6:8038.
- Saitsu H, Watanabe M, Akita T, Ohba C, Sugai K, Ong WP, Shiraishi H, Yuasa S, Matsumoto H, Beng KT, Saitoh S, Miyatake S, Nakashima M, Miyake N, Kato M, Fukuda A, Matsumoto N (2016) Impaired neuronal KCC2 function by biallelic SLC12A5 mutations in migrating focal seizures and severe developmental delay. Sci Rep 6:30072.
- Magloire V, Cornford J, Lieb A, Kullmann DM, Pavlov I (2019) KCC2 overexpression prevents the paradoxical seizure-promoting action of somatic inhibition. Nat Commun 10:1225.
- Coull JA, Boudreau D, Bachand K, Prescott SA, Nault F, Sík A, De Koninck P, De Koninck Y (2003) Trans-synaptic shift in anion gradient in spinal lamina I neurons as a mechanism of neuropathic pain. Nature 424:938-942.
- Lee HH, Deeb TZ, Walker JA, Davies PA, Moss SJ (2011) NMDA receptor activity downregulates KCC2 resulting in depolarizing GABAA receptor-mediated currents. Nat Neurosci 14:736-743.
- Medina I, Friedel P, Rivera C, Kahle KT, Kourdougli N, Uvarov P, Pellegrino C (2014) Current view on the functional regulation of the neuronal K(+)-Cl(-) cotransporter KCC2. Front Cell Neurosci 8:27.
- Hübner CA, Stein V, Hermans-Borgmeyer I, Meyer T, Ballanyi K, Jentsch TJ (2001) Disruption of KCC2 reveals an essential role of K-Cl cotransport already in early synaptic inhibition. Neuron 30:515-524.
- Zhu L, Lovinger D, Delpire E (2005) Cortical neurons lacking KCC2 expression show impaired regulation of intracellular chloride. J Neurophysiol 93:1557-1568.
- Blaesse P, Schmidt T (2015) K-Cl cotransporter KCC2--a moonlighting protein in excitatory and inhibitory synapse development and function. Pflugers Arch 467:615-624.
- Rivera C, Voipio J, Payne JA, Ruusuvuori E, Lahtinen H, Lamsa K, Pirvola U, Saarma M, Kaila K (1999) The K+/Cl- co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature 397:251-255.
- Li H, Khirug S, Cai C, Ludwig A, Blaesse P, Kolikova J, Afzalov R, Coleman SK, Lauri S, Airaksinen MS, Keinänen K, Khiroug L, Saarma M, Kaila K, Rivera C (2007) KCC2 interacts with the dendritic cytoskeleton to promote spine development. Neuron 56:1019-1033.
- Gauvain G, Chamma I, Chevy Q, Cabezas C, Irinopoulou T, Bodrug N, Carnaud M, Lévi S, Poncer JC (2011) The neuronal K-Cl cotransporter KCC2 influences postsynaptic AMPA receptor content and lateral diffusion in dendritic spines. Proc Natl Acad Sci U S A 108:15474-15479.
- Vanhatalo S, Palva JM, Andersson S, Rivera C, Voipio J, Kaila K (2005) Slow endogenous activity transients and developmental expression of K+-Cl- cotransporter 2 in the immature human cortex. Eur J Neurosci 22:2799-2804.
- Abbah J, Juliano SL (2014) Altered migratory behavior of interneurons in a model of cortical dysplasia: the influence of elevated GABAA activity. Cereb Cortex 24:2297-2308.
- Gagnon KB, Di Fulvio M (2013) A molecular analysis of the Na(+)-independent cation chloride cotransporters. Cell Physiol Biochem 32:14-31.
- Nakamura K, Moorhouse AJ, Cheung DL, Eto K, Takeda I, Rozenbroek PW, Nabekura J (2019) Overexpression of neuronal K+-Cl- co-transporter enhances dendritic spine plasticity and motor learning. J Physiol Sci 69:453-463.
- Pohl HR, Wheeler JS, Murray HE (2013) Sodium and potassium in health and disease. Met Ions Life Sci 13:29-47.
- Duménieu M, OuléM , Kreutz MR, Lopez-Rojas J (2017) The segregated expression of voltage-gated potassium and sodium channels in neuronal membranes: functional implications and regulatory mechanisms. Front Cell Neurosci 11:115.
- Huffaker SJ, Chen J, Nicodemus KK, Sambataro F, Yang F, Mattay V, Lipska BK, Hyde TM, Song J, Rujescu D, Giegling I, Mayilyan K, Proust MJ, Soghoyan A, Caforio G, Callicott JH, Bertolino A, Meyer-Lindenberg A, Chang J, Ji Y, Egan MF, Goldberg TE, Kleinman JE, Lu B, Weinberger DR (2009) A primate-specific, brain isoform of KCNH2 affects cortical physiology, cognition, neuronal repolarization and risk of schizophrenia. Nat Med 15:509-518.
- Heide J, Mann SA, Vandenberg JI (2012) The schizophrenia-associated Kv11.1-3.1 isoform results in reduced current accumulation during repetitive brief depolarizations. PLoS One 7:e45624.
- Blaesse P, Airaksinen MS, Rivera C, Kaila K (2009) Cation-chloride cotransporters and neuronal function. Neuron 61:820-838.
- Goutierre M, Al Awabdh S, Donneger F, François E, Gomez-Dominguez D, Irinopoulou T, Menendez de la Prida L, Poncer JC (2019) KCC2 regulates neuronal excitability and hippocampal activity via interaction with task-3 channels. Cell Rep 28:91-103.
- Spoljaric I, Spoljaric A, Mavrovic M, Seja P, Puskarjov M, Kaila K (2019) KCC2-mediated Cl- extrusion modulates spontaneous hippocampal network events in perinatal rats and mice. Cell Rep 26:1073-1081.
- Friedman RC, Farh KK, Burge CB, Bartel DP (2009) Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 19:92-105.
- MacDonald ML, Lamerdin J, Owens S, Keon BH, Bilter GK, Shang Z, Huang Z, Yu H, Dias J, Minami T, Michnick SW, Westwick JK (2006) Identifying off-target effects and hidden phenotypes of drugs in human cells. Nat Chem Biol 2:329-337.