Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2015; 24(2): 156-168
Published online June 30, 2015
https://doi.org/10.5607/en.2015.24.2.156
© The Korean Society for Brain and Neural Sciences
Chronic Antidepressant Treatment in Normal Mice Induces Anxiety and Impairs Stress-coping Ability
In-Sun Baek1,2#, Jin-Young Park1# and Pyung-Lim Han1,2*
Departments of 1Brain and Cognitive Sciences and 2Chemistry and Nano Science, Ewha Womans University, Seoul 120-750, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-3277-4130, FAX: 82-2-3277-3419
e-mail: plhan@ewha.ac.kr
#These authors contributed equally to this work.
Antidepressants are clinically used for patients with major depression. Antidepressant treatments in certain groups of patients are effective for relieving depression as well as anxiety disorder. However, it is not clearly known whether the use of current antidepressants in healthy persons is beneficial for upcoming depression- and anxiety-inducing life events. To address this question, normal mice were intraperitoneally administered with imipramine or fluoxetine for more than 2 weeks, and behaviors related to anxiety and depression were evaluated. Mice treated with imipramine or fluoxetine for more than 14 days exhibited significantly decreased immobility time in the forced swim test and tail suspension test, but these mice exhibited enhanced anxiety in several behavioral tests. Furthermore, chronic antidepressant treatments followed by sub-threshold level of stress in normal mice profoundly aggravated antidepressant-induced anxiety-like behaviors without further affecting depression-related behaviors. Chronic antidepressant treatments followed by sub-threshold level of stress produced swollen vesicles and ulcerations on the lips as well as a watery and inflammatory nose. Mice given chronic antidepressant treatments displayed intestinal abnormalities evidenced by a highly enlarged and inflamed small intestine full of defecation materials. These results suggest that chronic antidepressant treatment in normal mice provokes anxiety-like behaviors and impairs their stress-coping ability.
Keywords: Antidepressant, side effects, anxiety, stress-coping response
INTRODUCTION
Depression is one of the most prevalent psychiatric disorders and presents in patients as depressed mood, loss of interest, feelings of low self-worth, and diminished ability to experience pleasure [1,2]. Patients with depression are normally treated with antidepressants, which include classical monoamine modulators, such as tricyclic antidepressants (TCAs), monoamine oxidase inhibitors (MAOIs), selective serotonin reuptake inhibitors (SSRIs), and serotonin-norepinephrine reuptake inhibitors (SNRIs) [3,4]. However, current antidepressant medications are only effective in ~60% of patients [5]. Moreover, current antidepressants produce severe side effects that include dizziness, nausea, lethargy, headache, anxiety, agitation, and constipation [6,7,8,9,10].
Acute treatment with antidepressants in rodent models produces anxiogenic effects in the elevated plus maze test [11,12,13,14], light/dark box test [14], and social interaction test [15,16,17,18]. Conversely, other studies have reported that administration of acute antidepressants to normal animals produces opposing anxiolytic effects [19,20] or no effect [20,21]. Specifically, several studies have reported that chronic treatment of normal animals with antidepressants induces anxiogenic behaviors in the elevated plus maze, light/dark box test, novelty suppressed feeding test, open field test, and social interaction test [22,23,24,25], whereas a number of other studies have reported that chronic antidepressant treatments produce anxiolytic effects [11,12,26,27] or no effect at all [17,19,21,25,28,29]. Others have shown that treatment of normal animals with antidepressants does not change immobility time in the forced swim test [19,26,29,30], and may even increase immobility time in the forced swim test [31].
The conflicting results of antidepressant treatment effects in normal animals may be due in part to inconsistent experimental designs and skills in handling animals, antidepressant treatment procedures, and behavioral testing skills among different laboratories. Anxiety-like behaviors in animal models are easily induced by repeated stress [32,33], repetition of the behavioral tests [34,35,36], improper animal handling [37,38,39], and stressful housing conditions [40,41]. It is also unclear whether different antidepressants produce differential effects on depression- and anxiety-related behaviors [20]. Thus, the results of many previous studies are too complex to draw firm conclusions regarding anxiety. Nonetheless, in considering the effect of stress on anxiety, it is possible that antidepressant treatment during undefined subthreshold stress, which alone does not induce depression-like or anxiety-like behavioral changes, affected animal's anxiety states in previous studies. Therefore, chronic antidepressant treatment effects in normal animals need to be investigated in a more controlled manner in the context of depression, anxiety, and stress.
In the present study, we demonstrated that chronic treatments with imipramine or fluoxetine in mice induces anxiety-like behaviors, and that such treatments impaired animal's ability to cope with stress.
MATERIALS AND METHODS
Eight-week-old male C57BL/6J mice were purchased from Daehan BioLink, Inc (Eumsung, Chungbuk, Korea). Mice were housed in pairs into clear plastic cages in an environment in which temperature (23℃) and humidity (50~60%) were controlled on a 12-hr light/dark cycle (light on at 7 A.M.) with ad
Imipramine (Sigma-Aldrich, St. Louis, MO, USA) and fluoxetine (Sigma-Aldrich) were dissolved in 0.9% saline and injected intraperitoneally (i.p) daily at a dose of 20 mg/kg body weight in a total volume of 120 µl. Vehicle-treated animals received daily intraperitoneal injections of 120 µl saline.
Restraint was administered as described previously [32,42]. Mice were restrained for either 2 h or 6 h daily for 3 days as indicated. To administer restraint stress, animals were individually placed head-first into well-ventilated 50-ml conical tubes (SPL Lifesciences, Pocheon, Korea), plugged with a ~3-cm-long middle tube and finally the cap of the 50-ml tube. Mice could not freely move to the front or the reverse direction inside the tube. Restraint was delivered to animals beginning at 10 A.M. Control mice remained in their original cages with no disturbance. After 2 or 6 h-restraint treatment, the mice were returned to their home cage in which they were housed in pairs. Antidepressant or saline treated mice were administrated daily as indicated until the behavioral tests were finished.
Antidepressant- and stress-induced changes in pathological phenotypes were quantified as described previously [43]. Briefly, six days after the last stress session (day 17), the extent of antidepressant- and stress-induced pathological symptoms were assessed using rating scales ranging from 0 to 3. The scale for hair was as follows: 0, no observable change; +1 (weak), unkempt hairs on the head; +2 (moderate), unkempt and caked hairs on the head and neck; and +3 (severe), unkempt and caked hairs throughout the whole body. The scale for lips was as follows: 0, no observable change; +1 (weak), a sign of swelling on the lips; +2 (moderate), swelling, ulceration, and signs of scabs; and +3 (severe), inflammation and ulceration with noticeable scabs. Finally, the scale for nose was: 0, no observable change; +1 (weak), runny nose; +2 (moderate), runny nose with swellings and ulceration; and +3 (severe), runny nose with inflammation and ulceration along with respiratory difficulties.
Mice were brought to the testing room 30 min prior to the start of each behavioral test, and remained in the same room throughout the test. Behavioral assessments were performed using a computerized video-tracking system (SMART; Panlab S.I., Barcelona, Spain) for tracking mouse behavior or a webcam recording system, as previously described [32,44]. Behavioral tests were performed 1 h after the last antidepressant or vehicle injection.
The forced swim test (FST) was performed as described previously [32]. Briefly, mice were placed in a Plexiglas cylinder (height: 27 cm, diameter: 15 cm) containing water at 23℃ and a depth of 15 cm such that mice could neither escape nor touch the bottom. Mice were subjected to a 15-min pre-swim. After a 15-min pre-test, the animals were dried quickly with a towel and returned to their home cages. One day later, mice were forced to swim again for 6 min. The animals were habituated for the first 1 min and behavior was noted over the next 5 min. Immobility was defined as the summation of the time during which animals remained floating with all limbs motionless. This scoring method was applied by following the procedure described previously [45].
The tail suspension test (TST) was carried out as described previously [46] with minor modifications. Briefly, mice were individually suspended 50 cm above a table surface by fastening the tip (1 cm long) of their tail with tape. Immobility was defined as the total time during which all limbs were motionless. Immobility time was measured for 6 min-periods.
The sociability test (ST) was carried out by following the procedure described previously [47] with a minor modification. The sociability test was administered in the open field of a cream-colored FOAMAX (Expanded PVC; LG Ltd., Korea) chamber (45×45×40 cm). The testing chamber was lit with indirect illumination (50 lux). Mice were placed in the open field and left to explore the test arena. The time spent in the interaction zones and corner zone was assessed.
The open field test (OFT) was performed as described previously [32,48]. The apparatus of the open field test was built using cream-colored FOAMAX (45×45×40 cm). For the test, the intensity of the illumination in the open field chamber was adjusted to 80 lux by indirect illumination. Each mouse was placed individually at the center of the open field, and locomotion was recorded for the indicated period. Horizontal locomotor activity was assessed according to the total distance moved by the animal. The inner area, which was 30 percentage of the open field, was defined as the center zone.
The elevated plus maze (EPM) test was performed as described previously [32,48]. The elevated plus maze apparatus consisted of four arms (30×7 cm each) made of black-colored FOAMEX and placed at right angles to each other. Two of the arms had 20-cm high walls (enclosed arms), whereas the other two of the arms had no walls (open arms). The whole apparatus was elevated 50 cm above the test room floor. Illumination at the center of the apparatus was adjusted to 40 lux. For testing, each mouse was initially placed at the center of the platform and left to explore the arms for 5 min. The number of entries into the open and enclosed arms and the time spent in the each arm were recorded. The entry into any arm was scored as an event if the animal placed all four paws in the corresponding arm.
The novelty-suppressed feeding (NSF) test was administered as described previously [49] with minor modifications. Twenty-four hours before starting the test, mice were singly caged and food was removed from cage, but water was available ad
The marbles burying test (MBT) was administered as described previously [33,50] with minor modifications. Empty cages were filled with smooth bedding (JRS 3-4; J. Rettenmaier & Söhne, Rosenberg, Germany) to a height of 5 cm from the cage floor. Next, 12 identically shaped grass marbles were placed on top of the new bedding using equal spacing throughout the cage. Mice were individually tested by allowing them to freely explore the cage with marbles over a period of 30 min. Finally, the total number of buried marbles was counted. Buried marbles were defined as those in which the visible area of the marbles was less than 25% of the total marble surface.
The rota-rod test was performed as described previously [48]. The rota-rod consisted of a rotating cylinder (diameter 4.5 cm) with a speed controller. Mice were placed on top of the cylinder, which had a coarse surface to allow for a firm grip. The rota-rod was accelerated to a setting of 10 rpm, and maintained at the indicated speed for 3 min, and then the mice were subjected to tests at successively higher speeds. A cutoff time of 5 min and an inter-trial interval of 60 min were used. The time spent on the rod without falling down was measured.
Two-sample comparisons were performed using Student's
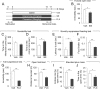
To understand whether chronic treatment with antidepressants affects depression- and anxiety-related behaviors, eight-week-old male C57BL/6J mice were intraperitoneally administered with imipramine (IMI, 20 mg/kg/day), a tricyclic antidepressant (TCA), for more than 14 consecutive days as described (Fig. 1A). Control animals received injections of saline (Veh) instead of the antidepressant for the same period of time. The behavioral states of mice on days 15~17 were examine. Chronic IMI-treated mice exhibited reduced immobility in the forced swim test (FST) [t(14)=2.332, p=0.0352] (Fig. 1B). The chronic IMI-treated mice interacted with a social target in the sociability test similar to Veh-treated control mice, and there was no statistical difference between chronic IMI-treated mice and Veh-treated control mice [t(24)=1.267, p=0.2173]. However, chronic IMI-treated mice spent more time in the corner zone than Veh-treated control mice [t(24)=2.529, p=0.0184] (Fig. 1C and D). In the novelty suppressed feeding (NSF) test, the IMI-treated mice showed increased latency to eating a food pellet placed at the center of the open field [t(12)=2.959, p=0.0119] (Fig. 1E), as well as reduced food consumption in a subsequent food consumption test [t(14)=4.525, p=0.0005] (Fig. 1F).
We examined whether chronic treatment of fluoxetine (FLX), a selective serotonin reuptake inhibitor (SSRI), produces similar behavioral effects as IMI. Normal C57BL/6J mice were treated with FLX (20 mg/kg/day) for more than 14 consecutive days as indicated above (Fig. 1A). Chronic FLX-treated mice showed reduced immobility in the tail suspension test (TST) [t(13)=7.898, p<0.0001] (Fig. 1G), and reduced locomotion in the open field test [t(12)=4.218, p=0.0012] (Fig. 1H). Chronic FLX-treated mice had reduced entry numbers to the open arms [t(13)=6.593, p<0.0001] and spent significantly less time in the open arms [t(11)=2.241, p=0.0466] in the elevated plus maze (EPM) test (Fig. 1I and J). These results suggested that chronic treatment with IMI or FLX in normal C57BL/6J mice suppresses depression-related behaviors, but induces enhanced anxiety compared with control animals.
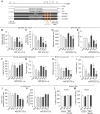
Next, we examined whether chronic antidepressant treatments in normal mice affect the stress coping abilities of mice in regulating anxiety-related behaviors. To address this, chronic IMI- or FLX-treated mice were exposed to restraint for 6 h daily for 3 days (6h×3d RST), according to the schedule depicted in Fig. 2A.
In the EPM test, chronic IMI treatment or 6h×3d RST treatment tended to decrease, but not significantly, entry numbers to the open arms and the amount of time spent in the open arms. Chronic IMI treatment followed by 6h×3d RST treatment significantly reduced entry numbers and the time spent in the open arms in the EPM test compared with chronic IMI treatment (Fig. 2B and C). Two-way ANOVA analysis detected the main effects of chronic IMI treatment [F(1,26)=17.62, p=0.0003 for entries; F(1,23)=8.242, p=0.0086 for time] and 6h×3d RST treatment [F(1,26)=14.01, p=0.0009 for entries; F(1,23)=9.456, p=0.0054 for time], whereas there was no effect of IMI×RST interaction [F(1,26)=3.297, p=0.0809 for entries; F(1,23)=1.667, p=0.2094 for time] (Fig. 2B and C). Chronic FLX treatment significantly decreased entries to the open arms and tended to reduce the amount of time spent in the open arms in the EPM test. The 6h×3d RST treatment effects were not significant in entries and the time as described above. Whereas chronic FLX treatment followed by 6h×3d RST treatment profoundly reduced open-arm entries and the time spent in the open arms compared with chronic FLX treatment (Fig. 2B and C). Two-way ANOVA analysis detected the main effect of chronic FLX treatment [F(1,27)=69.86, p<0.0001 for entries; F(1,24)=14.72, p=0.0008 for time], and 6h×3d RST treatment in open arm entries [F(1,27)=6.431, p=0.0173], but not in the time spent in the open arms [F(1,24)=3.359, p=0.0793]. There was no FLX×RST interaction [F(1,27)=0.3101, p=0.5822 for entries; F(1,24)=0.1739, p=0.6804 for time] (Fig. 2B and C).
Chronic IMI treatment in normal mice slightly reduced the total locomotion in the open field test (OFT), but did not change the amount of time spent in the center of the open field. The 6h×3d RST treatment in normal mice tended to increase the total locomotion and the amount of time spent in the center of the open field. Mice treated with chronic IMI followed by 6h×3d RST treatment did not significantly change the total locomotion and the amount of time spent in the center of the open field compared with chronic IMI treatment (Fig. 2D and E). Two-way ANOVA analysis detected the main effect of chronic IMI treatment in total locomotion [F(1,27)=6.152, p=0.0197], but not in time spent in the center [F(1,24)=2.234, p=0.1481]. There was no effect of 6h×3d RST treatment [F(1,27)=1.567, p=0.2213 for locomotion; F(1,24)=0.1301, p=0.7215 for center], and no IMI×RST interaction [F(1,27)=0.3161, p=0.5786 for locomotion; F(1,24)=0.1923, p=0.6649 for center] (Fig. 2D and E). Chronic FLX treatment in normal mice reduced total locomotion in the open field test and the amount of time spent in the center of the open field (Fig. 2D and E). Chronic FLX treatment followed by 6h×3d RST treatment did not change the reduced locomotion and the amount of time spent in the center of the open field compared with chronic FLX treatment (Fig. 2D and E). Two-way ANOVA analysis detected the main effect of chronic FLX treatment [F(1,29)=33.26, p<0.0001 for locomotion; F(1,27)=22.16, p<0.0001 for center], whereas there was no effect of 6h×3d RST treatment [F(1,29)=1.793, p=0.1910 for locomotion; F(1,27)=0.001775, p=0.9667 for center] and no FLX×RST interaction [F(1,29)=0.5954, p=0.4466 for locomotion; F(1,27)=0.00299, p=0.9568 for center] (Fig. 2D and E).
Chronic IMI or FLX treatment in normal mice significantly increased the latency for eating a food pellet placed in the center of the open field in the novelty suppressed feeding (NSF) test (Fig. 2F). In normal mice, the 6h×3d RST treatment tended to increase the latency in the NSF test. Chronic IMI or FLX treatment followed by 6h×3d RST treatment increased the latency, but the level was comparable to that by chronic IMI or FLX treatment (Fig. 2F). Two-way ANOVA analysis detected the main effects of chronic IMI or FLX treatment [F(1,31)=12.76, p=0.0012 for IMI; F(1,33)=17.84, p=0.0002 for FLX], but no or weak effect of 6h×3d RST treatment [F(1,31)=1.926, p=0.1751 for IMI; F(1,33)=4.134, p=0.0501 for FLX], and no antidepressant×RST interaction [F(1,31)=1.730, p=0.1981 for IMI×RST; F(1,33)=0.6891, p=0.4124 for FLX×RST] (Fig. 2F). In the subsequent assessment of food consumption, mice treated with chronic IMI showed slightly reduced food consumption, whereas mice treated with chronic IMI followed by 6h×3d RST treatment did not. Mice treated with chronic FLX exhibited reduced food consumption, and mice treated with chronic FLX followed by 6h×3d RST treatment also showed reduced food consumption, although the amount of food consumed was slightly enhanced after 6h×3d RST treatment (Fig. 2G). Two-way ANOVA analysis detected the main effect of chronic FLX treatment [F(1,33)=17.03, p=0.0002], but not IMI treatment [F(1,32)=4.014, p=0.0537]. There was no effect of 6h×3d RST treatment [F(1,32)=1.175, p=0.2865 for IMI; F(1,33)=0.5669, p=0.4568 for FLX], and no interaction between chronic IMI or FLX treatment and 6h×3d RST treatment [F(1,32)=1.037, p=0.3162 for IMI×RST; F(1,33)=0.4668, p=0.4992 for FLX×RST] (Fig. 2G).
Chronic IMI or FLX treatment in normal mice tended to suppress the number of marbles buried in the marble burying test (MBT). The 6h×3d RST treatment in normal mice decreased the number of marbles buried. Chronic FLX treatment followed by 6h×3d RST treatment suppressed the number of marbles buried to the level that was lower than that by chronic FLX treatment (Fig. 2H). Two-way ANOVA analysis detected the main effects of chronic IMI or FLX treatment [F(1,32)=6.273, p=0.0175 for IMI; F(1,33)=33.17, p<0.0001 for FLX], and the main effect of 6h×3d RST treatment in FLX group, but not in IMI group [F(1,32)=3.260, p=0.0804 for IMI; F(1,33)=8.234, p=0.0071 for FLX]. There were no antidepressant×RST interaction [F(1,32)=0.5734, p=0.4545 for IMI×RST; F(1,33)=0.02505, p=0.8752 for FLX×RST] (Fig. 2H).
Next, we examined whether chronic antidepressant treatment followed by short-term acute stress affects depression-related behaviors in normal mice (Fig. 2I, J and K). Chronic IMI treatment in normal mice significantly decreased immobility time in the tail suspension test (TST) (Fig. 2I). The 6h×3d RST treatment in normal mice did not significantly change immobility time in the TST. Chronic IMI treatment followed by 6h×3d RST treatment also decreased immobility time in the TST compared with chronic IMI treatment (Fig. 2I). Chronic FLX treatment in normal mice significantly reduced immobility time in the TST. Chronic FLX treatment followed by 6h×3d RST treatment suppressed immobility time in the TST compared with chronic FLX treatment (Fig. 2I). Two-way ANOVA analysis detected the main effects of chronic IMI or FLX treatment [F(1,28)=34.42, p<0.0001 for IMI; F(1,28)=683.2, p<0.0001 for FLX] and 6h×3d RST treatment [F(1,28)=8.421, p=0.0071 for IMI; F(1,28)=5.791, p=0.0230 for FLX]. There were no antidepressant×RST interaction [F(1,28)=0.8216, p=0.3724 for IMI×RST; F(1,28)=3.579, p=0.0689 for FLX×RST] (Fig. 2I).
Chronic IMI or FLX treatment in normal mice did not change the interaction time with a social target and the time spent in the corner zone in the sociability test. Likewise, the 6h×3d RST treatment did not change the interaction time with a social target and the time in the corner zone in the sociability test (Fig. 2J and K). Chronic IMI or FLX treatment followed by 6h×3d RST treatment tended to suppress the interaction time with a social target and tended to increase the time spent in the corner zone in the sociability test compared with chronic IMI or FLX treatment (Fig. 2J and K). Two-way ANOVA analysis showed no effect of chronic IMI treatment [F(1,25)=2.510, p=0.1257 for interaction zone; F(1,25)=3.354, p=0.0790 for corner zone] and chronic FLX treatment [F(1,25)=1.753, p=0.1975 for interaction zone; F(1,25)=2.478, p=0.1280 for corner zone] (Fig. 2J and K). There was the main effect of 6h×3d RST treatment in FLX group [F(1,25)=5.207, p=0.0313 for interaction zone; F(1,25)=4.348, p=0.0474 for corner zone], but not in IMI group [F(1,25)=2.344, p=0.1383 for interaction zone; F(1,25)=3.572, p=0.0704 for corner zone] (Fig. 2J and K). There were no antidepressant×RST interaction in IMI group [F(1,25)=0.3338, p=0.5686 for interaction zone; F(1,25)=1.201, p=0.2835 for corner zone] and in FLX group [F(1,25)=1.630, p=0.2135 for interaction zone; F(1,25)=1.629, p=0.2135 for corner zone] (Fig. 2J and K).
Chronic IMI treatment in normal mice did not change the riding performance on the rota-rod test. Likewise, the 6h×3d RST treatment did not change the performance on the rota-rod test. Chronic IMI treatment followed by 6h×3d RST treatment in normal mice also did not change the performance in the rota-rod test (Fig. 2L and M), indicating that chronic IMI treatment did not have reduced general motor activity. Two-way ANOVA analysis revealed no main effect of chronic IMI treatment [F(1,27)=0.01137, p=0.9159 for 10 rpm; F(1,28)=1.669, p=0.2070 for 15 rpm] and 6h×3d RST treatment [F(1,27)=1.811, p=0.1896 for 10 rpm; F(1,28)=2.026, p=0.1657 for 15 rpm], and no IMI×RST interaction [F(1,27)=0.5724, p=0.4559 for 10 rpm; F(1,28)=0.5578, p=0.4614 for 15 rpm] (Fig. 2L and M).
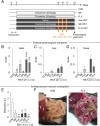
We next examined whether chronic antidepressant treatments in normal mice affected stress coping ability (Fig. 3). C57BL/6J mice that were exposed to restraint for 2 h daily for 3 days (2h×3d RST) actively cared for their coat states, thus maintaining well-kept hairs, in home cage returned after stress treatment procedure (Fig. 3A and B). On the contrary, chronic IMI or FLX treated mice neglected this behavior, such that their coat remained dirty and unkempt. Mice treated with chronic IMI followed by 2h×3d RST treatment showed more adverse coat state than chronic IMI group, thus indicative of more unhealthy overall appearance after 2h×3d RST treatment, whereas mice treated with chronic FLX followed by 2h×3d RST treatment looked unhealthy, but exhibited no more adverse change in coat state compared with chronic FLX group (Fig. 3B). Two-way ANOVA analysis detected the main effects of chronic IMI or FLX treatment [F(1,18)=30.77, p<0.0001 for IMI; F(1,17)=23.22, p=0.0002 for FLX] (Fig. 3B). There was the main effect of 2h×3d RST treatment in IMI, but not in FLX, group [F(1,18)=14.14, p=0.0014 for IMI; F(1,17)=0.513, p=0.4836 for FLX]. There was a significant interaction between IMI and RST, but not between FLX and RST [F(1,18)=10.01, p=0.0054 for IMI×RST; F(1,17)=0.06783, p=0.7976 for FLX×RST] (Fig. 3B).
Chronic IMI or FLX treatment in normal mice produced normal appearance of the lips and nose, although weak inflammatory signs on the lips and nose appeared in 1 animal among 5 animals. Chronic 2h×3d RST treatment produced no detectable sign on the lips and nose (Fig. 3C and D). Whereas chronic IMI or FLX treatment followed by 2h×3d RST induced the appearance of swollen vesicles and ulcerations on the lips (Fig. 3C) as well as watery and inflammatory noses in 5 out of 8 animals for IMI and 3 out of 7 animals for FLX, although the mean values of quantified symptoms were not statistically significant (Fig. 3D). Two-way ANOVA analysis showed no main effect of chronic IMI treatment [F(1,18)=0.9677, p=0.3383 for lips; F(1,18)=3.086, p=0.096 for nose] and chronic FLX treatment [F(1,17)=2.878, p=0.1081 for lips; F(1,17)=3.085, p=0.097 for nose]. There was no main effect of 2h×3d RST treatment in IMI group [F(1,18)=0.9677, p=0.3383 for lips; F(1,18)=3.086, p=0.096 for nose] and in FLX group [F(1,17)=0.9105, p=0.3534 for lips; F(1,17)=0.2455, p=0.6266 for nose]. There was no interaction between antidepressant and RST on lip symptom [F(1,18)=0.9677, p=0.3383 for IMI×RST; F(1,17)=0.9105, p=0.3534 for FLX×RST] and nose symptom [F(1,18)=3.086, p=0.096 for IMI×RST; F(1,17)=0.2455, p=0.6266 for FLX×RST] (Fig. 3C and D).
Chronic IMI or FLX treatment in normal mice decreased the number of defecations in the novel open field environment. The 6h×3d RST treatment in normal mice also decreased the number of defecations in the open field. Chronic IMI or FLX treatment followed by 6h×3d RST treatment in normal mice reduced the number of defecations in the open field compared with CON group (Fig. 3E). Two-way ANOVA analysis showed the main effect of chronic FLX treatment, but not chronic IMI treatment [F(1,32)=3.940, p=0.0558 for IMI; F(1,33)=15.16, p=0.0005 for FLX]. There was the main effect of 6h×3d RST treatment [F(1,32)=5.146, p=0.0302 for IMI; F(1,33)=6.767, p=0.0138 for FLX], and the significant antidepressant×RST interaction [F(1,32)=5.146, p=0.0302 for IMI×RST; F(1,33)=6.600, p=0.0149 for FLX×RST] (Fig. 3E).
Mice treated with chronic IMI or FLX followed by 6h×3d RST treatment usually had a large abdomen. Surgical opening of the abdomen revealed a highly enlarged and inflamed small intestine full of defecation materials (Fig. 3F).
The results of the present study showed that chronic treatment with imipramine or fluoxetine in normal mice produce enhanced anxiety in several behavioral tests. Moreover, chronic antidepressant treatment increased sensitivity of animals to subsequent sub-threshold levels of stress. These results raise several important questions concerning chronic treatment of imipramine or fluoxetine both in practice and research applications.
Major depression and anxiety disorders have high levels of comorbidity [51,52]. Among patients with major depression, 59.2% have anxiety disorders, while 45.5% of patients with anxiety disorders have major depression [51]. Mice treated with chronic stress show both depression- and anxiety-like behaviors in various animal models [25,32,33,53], and these anxiogenic phenotypes can be rescued by treatment with antidepressants [12,27]. Thus, depression is tightly associated with anxiety in certain patients with depression or in stress-induced depression animal models. On the contrary, the results of the present study suggest that certain depression- and anxiety-related behaviors within the same animals may express in opposite manners after chronic treatment with imipramine or fluoxetine.
The results of the present study indicate that chronic treatment with imipramine or fluoxetine in normal animals induces anxiety without affecting depressive-related behaviors (Fig. 1 and 2). Although the present study did not explore the detailed mechanism underlying antidepressant-induced enhanced anxiety, we can speculate upon following possibilities. Chronic imipramine or fluoxetine treatment may induce differential transcriptional changes in different brain regions, resulting in the suppression of depression-related behaviors while enhancing anxiety-related behaviors. Consistent with this possibility, chronic restraint stress produces differential gene expression profiles in the hippocampus and amygdala [32,54]. It is also possible that a single or a set of its related genes is induced by chronic antidepressant treatment commonly in different brain regions, which in turn facilitates suppression of depression-related behaviors while increasing anxiety-related behaviors. BDNF is one candidate gene which may produce such functions: (i) BDNF is induced by treatment with imipramine or fluoxetine in various regions of the brain, including the hippocampus [55,56] and
The results of the present study argue that chronic antidepressant treatment in normal mice deteriorate the animal's ability to withstand upcoming stress. Indeed, mice treated with antidepressants for 14 and more days showed weak or undetectable levels of physiological symptoms including changes in maintaining a clean coat state and normal immune responses as characterized by the appearance of the lips and nose (Fig. 3). On the other hand, animals treated with chronic antidepressants followed by a sub-threshold level of stress exhibited dirty and unkempt appearance of coat states (Fig. 3). These antidepressant-primed and stress-induced failures in maintaining proper coat state and immune responses that presented on the lips and nose, although the signs of lips and nose were subtle, were similar to those exhibited by AC5 KO mice exposed to short-term stress [43]. Specifically, AC5 KO mice exposed to restraint for 2 h daily for 5 days exhibit failure in keeping a proper coat state and have significant lips and nose signs. In addition, AC5 KO mice exposed to short-term stress have increased intestinal abnormalities as evidenced by a highly inflated and inflamed small intestine full of digested food (unpublished observations). In considering that AC5 KO mice displayed exaggerated stress responses [43], it will be interesting to investigate whether the hypothalamus-pituitary-adrenal gland (HPA) system is overactivated in mice undergoing chronic treatment with antidepressants.
In conclusion, we show that chronic treatment with imipramine or fluoxetine in normal mice results in a strong induction of anxiety, despite the fact that long-term treatment with these antidepressants confers anti-depressant-like effects. Chronic antidepressant treatments in normal mice also deteriorate the animal's stress coping ability, resulting is heightened sensitive to subsequent moderate stress. These results raise concerns regarding that long-term use of current anti-depressants and suggest that special considerations be made with respect to their use in practice and research applications.
{kind=link}
{kind=link}
{kind=link}
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders: DSM-5. 5th ed. Washington, D.C.: American Psychiatric Association, 2013.
- Williams JW, Noël PH, Cordes JA, Ramirez G, Pignone M. Is this patient clinically depressed?. JAMA 2002;287:1160-1170.

- Holtzheimer PE, Nemeroff CB. Advances in the treatment of depression. NeuroRx 2006;3:42-56.
- Olfson M, Marcus SC. National patterns in antidepressant medication treatment. Arch Gen Psychiatry 2009;66:848-856.
- Souery D, Papakostas GI, Trivedi MH. Treatment-resistant depression. J Clin Psychiatry 2006;67:16-22.
- Healy D. The psychopharmacologists II: interviews by Dr David Healy. London: Arnold, 1998.
- Cascade E, Kalali AH, Kennedy SH. Real-world data on SSRI antidepressant side effects. Psychiatry (Edgmont) 2009;6:16-18.
- Brambilla P, Cipriani A, Hotopf M, Barbui C. Side-effect profile of fluoxetine in comparison with other SSRIs, tricyclic and newer antidepressants: a meta-analysis of clinical trial data. Pharmacopsychiatry 2005;38:69-77.
- Furukawa TA, McGuire H, Barbui C. Meta-analysis of effects and side effects of low dosage tricyclic antidepressants in depression: systematic review. BMJ 2002;325:991.
- Sinclair LI, Christmas DM, Hood SD, Potokar JP, Robertson A, Isaac A, Srivastava S, Nutt DJ, Davies SJ. Antidepressant-induced jitteriness/anxiety syndrome: systematic review. Br J Psychiatry 2009;194:483-490.
- Silva RC, Brandão ML. Acute and chronic effects of gepirone and fluoxetine in rats tested in the elevated plus-maze: an ethological analysis. Pharmacol Biochem Behav 2000;65:209-216.
- Mombereau C, Gur TL, Onksen J, Blendy JA. Differential effects of acute and repeated citalopram in mouse models of anxiety and depression. Int J Neuropsychopharmacol 2010;13:321-334.
- Drapier D, Bentué-Ferrer D, Laviolle B, Millet B, Allain H, Bourin M, Reymann JM. Effects of acute fluoxetine, paroxetine and desipramine on rats tested on the elevated plus-maze. Behav Brain Res 2007;176:202-209.
- Griebel G, Moreau JL, Jenck F, Misslin R, Martin JR. Acute and chronic treatment with 5-HT reuptake inhibitors differentially modulate emotional responses in anxiety models in rodents. Psychopharmacology (Berl) 1994;113:463-470.
- Bagdy G, Graf M, Anheuer ZE, Modos EA, Kantor S. Anxiety-like effects induced by acute fluoxetine, sertraline or m-CPP treatment are reversed by pretreatment with the 5-HT2C receptor antagonist SB-242084 but not the 5-HT1A receptor antagonist WAY-100635. Int J Neuropsychopharmacol 2001;4:399-408.
- To CT, Bagdy G. Anxiogenic effect of central CCK administration is attenuated by chronic fluoxetine or ipsapirone treatment. Neuropharmacology 1999;38:279-282.
- File SE, Ouagazzal AM, Gonzalez LE, Overstreet DH. Chronic fluoxetine in tests of anxiety in rat lines selectively bred for differential 5-HT1A receptor function. Pharmacol Biochem Behav 1999;62:695-701.
- To CT, Anheuer ZE, Bagdy G. Effects of acute and chronic fluoxetine treatment of CRH-induced anxiety. Neuroreport 1999;10:553-555.
- Griebel G, Cohen C, Perrault G, Sanger DJ. Behavioral effects of acute and chronic fluoxetine in Wistar-Kyoto rats. Physiol Behav 1999;67:315-320.
- Sánchez C, Meier E. Behavioral profiles of SSRIs in animal models of depression, anxiety and aggression. Are they all alike?. Psychopharmacology (Berl) 1997;129:197-205.
- Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, Graham D, Tsankova NM, Bolanos CA, Rios M, Monteggia LM, Self DW, Nestler EJ. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006;311:864-868.
- Bondi CO, Rodriguez G, Gould GG, Frazer A, Morilak DA. Chronic unpredictable stress induces a cognitive deficit and anxiety-like behavior in rats that is prevented by chronic antidepressant drug treatment. Neuropsychopharmacology 2008;33:320-331.
- Durand M, Berton O, Aguerre S, Edno L, Combourieu I, Mormède P, Chaouloff F. Effects of repeated fluoxetine on anxiety-related behaviours, central serotonergic systems, and the corticotropic axis axis in SHR and WKY rats. Neuropharmacology 1999;38:893-907.
- Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, Herrera DG, Toth M, Yang C, McEwen BS, Hempstead BL, Lee FS. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 2006;314:140-143.
- Venzala E, García-García AL, Elizalde N, Delagrange P, Tordera RM. Chronic social defeat stress model: behavioral features, antidepressant action, and interaction with biological risk factors. Psychopharmacology (Berl) 2012;224:313-325.
- Thompson MR, Li KM, Clemens KJ, Gurtman CG, Hunt GE, Cornish JL, McGregor IS. Chronic fluoxetine treatment partly attenuates the long-term anxiety and depressive symptoms induced by MDMA ('Ecstasy') in rats. Neuropsychopharmacology 2004;29:694-704.
- Dulawa SC, Holick KA, Gundersen B, Hen R. Effects of chronic fluoxetine in animal models of anxiety and depression. Neuropsychopharmacology 2004;29:1321-1330.
- Oh JE, Zupan B, Gross S, Toth M. Paradoxical anxiogenic response of juvenile mice to fluoxetine. Neuropsychopharmacology 2009;34:2197-2207.
- Wagner KV, Marinescu D, Hartmann J, Wang XD, Labermaier C, Scharf SH, Liebl C, Uhr M, Holsboer F, Müller MB, Schmidt MV. Differences in FKBP51 regulation following chronic social defeat stress correlate with individual stress sensitivity: influence of paroxetine treatment. Neuropsychopharmacology 2012;37:2797-2808.
- Balsevich G, Namendorf C, Gerlach T, Uhr M, Schmidt MV. The bio-distribution of the antidepressant clomipramine is modulated by chronic stress in mice: effects on behavior. Front Behav Neurosci 2014;8:445.
- Marrocco J, Reynaert ML, Gatta E, Gabriel C, Mocaër E, Di Prisco S, Merega E, Pittaluga A, Nicoletti F, Maccari S, Morley-Fletcher S, Mairesse J. The effects of antidepressant treatment in prenatally stressed rats support the glutamatergic hypothesis of stress-related disorders. J Neurosci 2014;34:2015-2024.
- Kim KS, Han PL. Optimization of chronic stress paradigms using anxiety- and depression-like behavioral parameters. J Neurosci Res 2006;83:497-507.
- Kim KS, Kwon HJ, Baek IS, Han PL. Repeated short-term (2h×14d) emotional stress induces lasting depression-like behavior in mice. Exp Neurobiol 2012;21:16-22.
- Võikar V, Vasar E, Rauvala H. Behavioral alterations induced by repeated testing in C57BL/6J and 129S2/Sv mice: implications for phenotyping screens. Genes Brain Behav 2004;3:27-38.
- Almeida SS, Garcia RA, de Oliveira LM. Effects of early protein malnutrition and repeated testing upon locomotor and exploratory behaviors in the elevated plus-maze. Physiol Behav 1993;54:749-752.
- Cook MN, Crounse M, Flaherty L. Anxiety in the elevated zero-maze is augmented in mice after repeated daily exposure. Behav Genet 2002;32:113-118.
- Hurst JL, West RS. Taming anxiety in laboratory mice. Nat Methods 2010;7:825-826.
- Balcombe JP, Barnard ND, Sandusky C. Laboratory routines cause animal stress. Contemp Top Lab Anim Sci 2004;43:42-51.
- Schmitt U, Hiemke C. Strain differences in open-field and elevated plus-maze behavior of rats without and with pretest handling. Pharmacol Biochem Behav 1998;59:807-811.
- Liu X, Wu R, Tai F, Ma L, Wei B, Yang X, Zhang X, Jia R. Effects of group housing on stress induced emotional and neuroendocrine alterations. Brain Res 2013;1502:71-80.
- Rodgers RJ, Dalvi A. Anxiety, defence and the elevated plus-maze. Neurosci Biobehav Rev 1997;21:801-810.
- Park JY, Kim TK, Choi J, Lee JE, Kim H, Lee EH, Han PL. Implementation of a two-dimensional behavior matrix to distinguish individuals with differential depression states in a rodent model of depression. Exp Neurobiol 2014;23:215-223.
- Kim KS, Han PL. Mice lacking adenylyl cyclase-5 cope badly with repeated restraint stress. J Neurosci Res 2009;87:2983-2993.
- Lee KW, Lee SH, Kim H, Song JS, Yang SD, Paik SG, Han PL. Progressive cognitive impairment and anxiety induction in the absence of plaque deposition in C57BL/6 inbred mice expressing transgenic amyloid precursor protein. J Neurosci Res 2004;76:572-580.
- Porsolt RD, Bertin A, Jalfre M. Behavioral despair in mice: a primary screening test for antidepressants. Arch Int Pharmacodyn Ther 1977;229:327-336.
- Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl) 1985;85:367-370.
- Salchner P, Singewald N. Neuroanatomical substrates involved in the anxiogenic-like effect of acute fluoxetine treatment. Neuropharmacology 2002;43:1238-1248.
- Lee KW, Im JY, Song JS, Lee SH, Lee HJ, Ha HY, Koh JY, Gwag BJ, Yang SD, Paik SG, Han PL. Progressive neuronal loss and behavioral impairments of transgenic C57BL/6 inbred mice expressing the carboxy terminus of amyloid precursor protein. Neurobiol Dis 2006;22:10-24.
- Lira A, Zhou M, Castanon N, Ansorge MS, Gordon JA, Francis JH, Bradley-Moore M, Lira J, Underwood MD, Arango V, Kung HF, Hofer MA, Hen R, Gingrich JA. Altered depression-related behaviors and functional changes in the dorsal raphe nucleus of serotonin transporter-deficient mice. Biol Psychiatry 2003;54:960-971.
- Deacon RM. Digging and marble burying in mice: simple methods for in vivo identification of biological impacts. Nat Protoc 2006;1:122-124.
- Kessler RC, Berglund P, Demler O, Jin R, Koretz D, Merikangas KR, Rush AJ, Walters EE, Wang PS, National Comorbidity Survey Replication. The epidemiology of major depressive disorder: results from the National Comorbidity Survey Replication (NCS-R). JAMA 2003;289:3095-3105.
- Kessler RC, Chiu WT, Demler O, Merikangas KR, Walters EE. Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry 2005;62:617-627.
- Zhu S, Wang J, Zhang Y, Li V, Kong J, He J, Li XM. Unpredictable chronic mild stress induces anxiety and depression-like behaviors and inactivates AMP-activated protein kinase in mice. Brain Res 2014;1576:81-90.
- Kim TK, Kim JE, Park JY, Lee JE, Choi J, Kim H, Lee EH, Kim SW, Lee JK, Kang HS, Han PL. Antidepressant effects of exercise are produced via suppression of hypocretin/orexin and melanin-concentrating hormone in the basolateral amygdala. Neurobiol Dis 2015;79:59-69.
- Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL, Nestler EJ. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat Neurosci 2006;9:519-525.
- Baj G, D'Alessandro V, Musazzi L, Mallei A, Sartori CR, Sciancalepore M, Tardito D, Langone F, Popoli M, Tongiorgi E. Physical exercise and antidepressants enhance BDNF targeting in hippocampal CA3 dendrites: further evidence of a spatial code for BDNF splice variants. Neuropsychopharmacology 2012;37:1600-1611.
- Seo MK, Lee CH, Cho HY, Lee JG, Lee BJ, Kim JE, Seol W, Kim YH, Park SW. Effects of antidepressant drugs on synaptic protein levels and dendritic outgrowth in hippocampal neuronal cultures. Neuropharmacology 2014;79:222-233.
- Taliaz D, Loya A, Gersner R, Haramati S, Chen A, Zangen A. Resilience to chronic stress is mediated by hippocampal brain-derived neurotrophic factor. J Neurosci 2011;31:4475-4483.
- Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ, Laplant Q, Graham A, Lutter M, Lagace DC, Ghose S, Reister R, Tannous P, Green TA, Neve RL, Chakravarty S, Kumar A, Eisch AJ, Self DW, Lee FS, Tamminga CA, Cooper DC, Gershenfeld HK, Nestler EJ. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007;131:391-404.
- Fanous S, Hammer RP, Nikulina EM. Short- and long-term effects of intermittent social defeat stress on brain-derived neurotrophic factor expression in mesocorticolimbic brain regions. Neuroscience 2010;167:598-607.
- Lakshminarasimhan H, Chattarji S. Stress leads to contrasting effects on the levels of brain derived neurotrophic factor in the hippocampus and amygdala. PLoS One 2012;7:e30481.
- Yu H, Wang DD, Wang Y, Liu T, Lee FS, Chen ZY. Variant brain-derived neurotrophic factor Val66Met polymorphism alters vulnerability to stress and response to antidepressants. J Neurosci 2012;32:4092-4101.
- Eisch AJ, Bolaños CA, de Wit J, Simonak RD, Pudiak CM, Barrot M, Verhaagen J, Nestler EJ. Brain-derived neurotrophic factor in the ventral midbrain-nucleus accumbens pathway: a role in depression. Biol Psychiatry 2003;54:994-1005.
- Pandey SC, Zhang H, Roy A, Misra K. Central and medial amygdaloid brain-derived neurotrophic factor signaling plays a critical role in alcohol-drinking and anxiety-like behaviors. J Neurosci 2006;26:8320-8331.
- Govindarajan A, Rao BS, Nair D, Trinh M, Mawjee N, Tonegawa S, Chattarji S. Transgenic brain-derived neurotrophic factor expression causes both anxiogenic and antidepressant effects. Proc Natl Acad Sci U S A 2006;103:13208-13213.
- MacQueen GM, Ramakrishnan K, Croll SD, Siuciak JA, Yu G, Young LT, Fahnestock M. Performance of heterozygous brain-derived neurotrophic factor knockout mice on behavioral analogues of anxiety, nociception, and depression. Behav Neurosci 2001;115:1145-1153.
- Saarelainen T, Hendolin P, Lucas G, Koponen E, Sairanen M, MacDonald E, Agerman K, Haapasalo A, Nawa H, Aloyz R, Ernfors P, Castren E. Activation of the TrkB neurotrophin receptor is induced by antidepressant drugs and is required for antidepressant-induced behavioral effects. J Neurosci 2003;23:349-357.
- Chourbaji S, Hellweg R, Brandis D, Zorner B, Zacher C, Lang UE, Henn FA, Hörtnagl H, Gass P. Mice with reduced brain-derived neurotrophic factor expression show decreased choline acetyltransferase activity, but regular brain monoamine levels and unaltered emotional behavior. Brain Res Mol Brain Res 2004;121:28-36.
- Ibarguen-Vargas Y, Surget A, Vourc'h P, Leman S, Andres CR, Gardier AM, Belzung C. Deficit in BDNF does not increase vulnerability to stress but dampens antidepressant-like effects in the unpredictable chronic mild stress. Behav Brain Res 2009;202:245-251.
- Lindholm JS, Castrén E. Mice with altered BDNF signaling as models for mood disorders and antidepressant effects. Front Behav Neurosci 2014;8:143.