Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2010; 19(2): 106-113
Published online September 30, 2010
https://doi.org/10.5607/en.2010.19.2.106
© The Korean Society for Brain and Neural Sciences
Organotypic Spinal Cord Slice Culture to Study Neural Stem/Progenitor Cell Microenvironment in the Injured Spinal Cord
Hyuk Min Kim1†, Hong Jun Lee2†, Man Young Lee1, Seung U. Kim2,3 and Byung Gon Kim1*
1Brain Disease Research Center, Institute for Medical Sciences, and Department of Neurology, Ajou University School of Medicine, Suwon 442-721, Korea, 2Medical Research Institute, Chungang University School of Medicine, Seoul 156-756, Korea, 3Department of Neurology, University of British Columbia, Vancouver, BC V6T 2B5, Canada
Correspondence to: *To whom correspondence should be addressed.
†The first two authors equally contributed to this work
TEL: 82-31-219-4495, FAX: 82-31-219-4530
e-mail: kimbg@ajou.ac.kr
The molecular microenvironment of the injured spinal cord does not support survival and differentiation of either grafted or endogenous NSCs, restricting the effectiveness of the NSC-based cell replacement strategy. Studying the biology of NSCs in
Keywords: spinal cord injury, organotypic slice culture, neural stem cells, hypoxia, inflammatory cytokine
INTRODUCTION
Traumatic injuries to the spinal cord frequently leave permanent neurological disabilities to the victims and impose enormous economic burdens on the families and society, yet there is no single effective therapeutic option to improve functional recovery. Recent studies have shown promises that cellular replacement either by transplantation of neural stem or progenitor cells (NSC) or mobilization of endogenous NSCs could be an effective therapeutic option (Rossignol et al., 2007; Meletis et al., 2008; Cao et al., 2010; Tetzlaff et al., 2010). However, inhospitable microenvironment of injured spinal cord has been shown to limit survival and differentiation of either grafted or endogenous NSCs (Monje et al., 2002; Snyder and Park, 2002; Ishii et al., 2006; Kim et al., 2007). For example, survival of NSC continually dropped after transplantation (Okada et al., 2005; Lee et al., 2009). Moreover, differentiation of NSCs into neuronal or oligodendroglial lineages in the injured spinal cord is significantly hampered (Cao et al., 2001; Cao et al., 2002). Therefore, modification of the inhospitable microenvironment would greatly improve the efficacy of NSC transplantation approach for spinal cord injury.
Since the complexity of cellular and molecular composition can hardly be modeled in dissociated cell culture system, studying potential factors that regulate the microenvironment would necessitate
MATERIALS AND METHODS
As an exogenous source of NSCs, we used immortalized human neural stem cell (NSC) line, which has been widely employed in various animal models of CNS diseases (Jeong et al., 2003; Meng et al., 2003; Yasuhara et al., 2006; Lee et al., 2007; Hwang et al., 2009). Preparation and culture of this NSC line has been reported in detail elsewhere (Lee et al., 2009). Briefly, the human NSC line was generated by transducing dissociated cells of fetal human telencephalon tissues (at 14 weeks gestation) by replication incompetent retroviral vector containing v-myc (Flax et al., 1998; Kim, 2004). The permission to use the fetal tissues was granted by the Clinical Research Screening Committee involving Human Subjects of the University of British Columbia, and the fetal tissues were obtained from the Anatomical Pathology Department of Vancouver General Hospital. Cryopreserved NSCs were thawed and cultured in Dulbecco's modified Eagle medium (DMEM; HyClone, Logan, UT, USA) with high glucose supplemented with 5% fetal bovine serum (FBS) and 20 mg/ml gentamicin (Sigma, St Louis, MO, USA) for at least three days before cell seeding. We also generated a NSC line overexpressing green fluorescent protein (GFP) by transducing the human NSC line by retrovirus encoding GFP.
Organotypic spinal cord slice cultures were prepared according to the standard interface method (Stoppini et al., 1991). After decapitation, the brain was removed, and the entire spinal cord block was dissected from P5-7 Sprague Dawley rats through an opening in the ventral side of the spine. Axial slices of the cervical and lumbar cord were dissected and transversely sliced into a 350 um thickness on a McILWAIN tissue chopper (The Mickle Laboratory Engineering Co., Guildford, UK) in sterile Gey's balanced salt solution (Sigma-Aldrich). The slices were then carefully separated with two pairs of fine forceps and transferred to sterile, 30 mm diameter Millipore Milicell-CM (0.4 µm; Millipore, Bedford, MA) culture plate insert, using a glass Pasteur pipette. Five or six randomly selected slices that looked apparently intact and undamaged were transferred and placed on each insert. The inserts were placed in 35 mm diameter culture wells (six well culture trays; BD Falcon, Franklin Lakes, NJ). Cultures were maintained in 1 ml of the serum-based medium containing 50% Basal Medial Eagle (Sigma-Aldrich), 25% Hank's Balanced Salt Solution (GIBCO), 2.2 g glucose, 1 mM GlutaMAX-I supplement (Invitrogen, Carlsbad, CA), and 20% FBS. Culture plates were incubated at 37℃ in a 5% CO2-95% O2 humidified incubator. Culture medium was changed 4 hours after harvesting and then twice per week. The level of the medium was adjusted to slightly below the surface of the slices in order to provide a sufficient supply of the culture medium and mixed gases. Cell seeding was performed 7 days after initial slice preparation. The human NSCs were trypsinized just before seeding, and a total of 1,000 cell/1µl cells for each slice were seeded using a glass micropipette. Special care was taken to avoid touching slices with micropipette. One day after seeding, the culture medium was changed. To induce differentiation of NSCs grown on top of slices, the FBS concentration was lowered to 5%. To identify seeded NSCs, cells were prelabeled with BrdU or DiI. For BrdU prelabeling, cells were treated with 2 µM BrdU dissolved in culture media for 24 hours prior to harvesting for transplantation. The human NSCs were labeled Vybrant™DiI (Molecular Probe) according to the manufacturer's instruction. To mark endogenous proliferating neural progenitor cells, BrdU at a concentration of 1.0µM was added to the culture media one day before fixation (24 hours incubation). IL-1β (R&D systems, Minneapolis, MN) was added to the media at a concentration of 20 ng/ml for three days before fixation.
To mimic secondary injury process after spinal cord injury, cultured spinal cord slices were exposed to a hypoxic chamber (Forma Scientific, Marietta, OH). Glucose-free medium DMEM was saturated with nitrogen gas mixture (95% N2, and 5% CO2) for 40 min to obtain an O2 gas pressure close to zero, as measured by a dip-type O2 microelectrode. After saturation, the inserts with spinal cord slices were placed in 1 ml of saturated glucose-free medium DMEM and then maintained at 37℃ in a N2 saturated environment. Therefore, the cultured slices were challenged by aglycemic hypoxic stress. After 40 minutes in the chamber, the inserts were moved to the fresh culture medium and atmosphere with 5% CO2-95% O2. One day after, NSCs were seeded as described above.
Slices were washed in PBS and fixed in 4% paraformaldehyde for 5 min. Slice was excised from the culture insert together with the attached membrane, and each slice is transferred to a 24 well plate. The slices were permeabilized and blocked by 0.5% triton with 10% goat serum for 2 hours. Then, slices were incubated overnight with primary antibodies at 4℃ or 2 hours at room temperature. We used polyclonal NG2 antibody (1:1,000; Millipore, Bedford, MA) as a marker for oligodendrocyte progenitors, polyclonal GFAP antibody (1:500; Dako, Carpinteria, CA) for astrocytes, Tuj1 (1:500; Millipore, Bedford, MA) for immature neurons, CD11b (1:300: Abcam, Cambridge, UK) for resident microglia, and BrdU (1:500; Serotec, Oxford, UK) for a marker of proliferating NSCs. For BrdU staining, DNA denaturation was achieved by treatment with 2 M HCl at room temperature for 60 min followed by incubation for 30 min with 0.1 M Borate solution (Sigma-Aldrich). After thorough rinsing, slices were incubated by rat IgG secondary antibody tagged with Alexa Fluor 594 or 488 (Molecular Probes, Eugene, OR) for 1 hour at room temperature to visualize antigen-antibody complex.
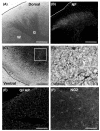
We first characterized the histological architecture of cultured spinal cord slices. When the cultured slice was view with the transmitted light, the gray matter was apparently distinguished from the surrounding white matter (Fig. 1A). Neurofilament staining revealed clear margin of the dorsal horn where profuse axons and scattered neuronal cell bodies were observed (Fig. 1B). In contrast, axonal fibers in the white matter were sparsely observed, suggesting that axons in the white matter tract underwent some degree of degeneration after disconnected from the cell bodies. In the ventral horn, large neurons suggestive of spinal motor neurons were observed (Fig. 1C, D). They grew long neurites within the slice indicating that the neurons were healthy and made connections with different neurons in the slice. We also characterized glial cells in the spinal cord slices. GFAP staining showed a large number of astrocytes located in both the white and gray matter (Fig. 1E). Numerous oligodendrocyte progenitors expressing NG2 proteoglycan (Dawson et al., 2000) were also observed. However, CD11b (OX42) staining did not show apparent microglial cells in the slice (data not shown), suggesting that no macrophages migrate to the spinal cord to become resident microglial cells before postnatal day 5. Together, these results showed that the spinal cord slices maintain characteristics of the cellular and tissue architecture of the
To mimic NSC grafting into the spinal cord, we seeded NSCs on top of cultured spinal cord slices using the Hamilton syringe at between 7 to 10 days after initial culture. The seeded hNSCs on spinal cord slice culture were prelabeled by DiI or BrdU. In some experiments, we used GFP expressing hNSCs to identify the seeded cells. Many of the cells survived the seeding procedure and were identified up to 4 weeks after seeding (Fig. 2A~D), the last time point we measured. They were dispersed throughout the surface of the spinal cord slice, but many of them were found at or near the parts of the slice where they were dislodged from the Hamilton syringe (Fig. 2B). Dispersing cells did not show any preference to either the white or gray matter. These findings indicated that NSCs can be cultured on top of cultured spinal cord slices, providing an opportunity to examine biological behavior of NSCs in an environment which closely mimics
The spinal cord has been regarded as nonneurogenic. When NSCs collected from spinal cord were transplanted into spinal cord, they could not differentiate into neurons, whereas the same NSCs differentiated neurons when they were transplanted into the brain (Shihabuddin et al., 2000). We examined differentiation of NSCs co-cultured on spinal cord slices. NSCs did not express neuronal marker Tuj-1 (Fig. 3A). Addition of retinoic acid into the culture medium did not increase Tuj-1 expression (data not shown). Some NSCs (20% of BrdU+ cells) were colocalized with GFAP, indicating differentiation into astrocytic lineage (Fig. 3B). In addition, they were able to differentiate into NG2 positive oligodendrocytic lineage cells (26% of BrdU+ cells; Fig. 3C). Therefore, the microenvironment created by cultured spinal cord tissue does not seem to be conducive to neuronal differentiation of NSCs, lending a support to the notion that spinal cord is non-neurogenic.
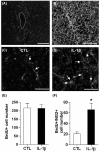
Endogenous glial progenitor cells are present in adult spinal cord and increase their number in response to spinal cord injury (Horner et al., 2000; McTigue et al., 2001). It is possible that they can differentiate into mature oligodendrocytes that may participate in spontaneous remyelination process (Yang et al., 2006), although the extent of remyelination is often limited. To recapitulate the glial progenitor cells in a complex environment, we marked proliferating neural progenitor cells by BrdU incorporation (Fig. 4A). As expected from the
We found that in organotypic spinal cord slices, regional specificity such as gray and white matter is conserved and cellular diversity and/or complexity is maintained encompassing neurons and glial cells. The fact that the cultured slices retain major characteristics of
It has been increasingly clear that adult CNS, especially diseased CNS, is not always favorable to the integration NSCs with host tissue (Bjorklund and Lindvall, 2000; Snyder and Park, 2002; Okano et al., 2003). Injured CNS microenvironment considerably limits the survival of grafted cells (Emgard et al., 2003; Bakshi et al., 2005; Lee et al., 2009), which may pose a significant hurdle to be overcome before NSC transplantation strategy is applied to human patients. Traumatic spinal cord injuries are usually complicated by a breakdown of blood supply leading to tissue ischemia and hypoxia (Chu et al., 2002). As an example of altering the microenvironment in a manner similar to the spinal cord injury, the cultured spinal cord slices were pre-exposed to a hypoxic (aglycemic as well) chamber before the NSCs were seeded on the them. Although hypoxic injury is not supposed to replicate all the changes related to traumatic injuries, we found that exposure to hypoxic chamber for 40 minutes did make a difference in the survival of seeded NSCs on the slices. It is assumed that hypoxic conditions altered the environment of spinal cord slices to become more inhospitable for NSCs to survive. Therefore, the hypoxic condition used in this experiment can be used to screen potential factors or small molecules that regulate the survival of grafted NSCs in the injured spinal cord.
After spinal cord injury, demyelination of spared white matter significantly hampers spontaneous function recovery (Kim et al., 2007). Therefore, preventing demyelination or promoting remyelination is one of the key strategies to improve function outcomes after spinal cord injury (McDonald and Belegu, 2006). Modifying the microenvironment of the injured spinal cord may improve the extent of oligodendrogenesis and ultimately promote remyelination. We tested whether inflammatory molecules can affect the fate of glial progenitors, especially oligodendrocytic lineage, in the cultured spinal cord slice. The cultured spinal cord slices closely recapitulated
To summarize, the current study established the utility of the organotypic spinal cord slices to study neural stem/progenitor cell microenvironment in the injured spinal cord. Co-culture of exogenous NSCs was feasible mimicking post-graft environment. Exposure of the cultured slices to hypoxic chamber mimicked the post-injury environment in that the survival of seeded NSCs was reduced. Cultured spinal cord slices retained the non-neurogenic characteristics of


{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Ankeny DP, Popovich PG. Mechanisms and implications of adaptive immune responses after traumatic spinal cord injury. Neuroscience 2009;158:1112-1121.

- Bakshi A, Keck CA, Koshkin VS, LeBold DG, Siman R, Snyder EY, McIntosh TK. Caspase-mediated cell death predominates following engraftment of neural progenitor cells into traumatically injured rat brain. Brain Research 2005;1065:8-19.
- Bjorklund A, Lindvall O. Cell replacement therapies for central nervous system disorders. Nat Neurosci 2000;3:537-544.
- Cao Q, He Q, Wang Y, Cheng X, Howard RM, Zhang Y, DeVries WH, Shields CB, Magnuson DS, Xu XM, Kim DH, Whittemore SR. Transplantation of ciliary neurotrophic factor-expressing adult oligodendrocyte precursor cells promotes remyelination and functional recovery after spinal cord injury. J Neurosci 2010;30:2989-3001.
- Cao QL, Howard RM, Dennison JB, Whittemore SR. Differentiation of engrafted neuronal-restricted precursor cells is inhibited in the traumatically injured spinal cord. Exp Neurol 2002;177:349-359.
- Cao QL, Zhang YP, Howard RM, Walters WM, Tsoulfas P, Whittemore SR. Pluripotent stem cells engrafted into the normal or lesioned adult rat spinal cord are restricted to a glial lineage. Exp Neurol 2001;167:48-58.
- Chu D, Qiu J, Grafe M, Fabian R, Kent TA, Rassin D, Nesic O, Werrbach-Perez K, Perez-Polo R. Delayed cell death signaling in traumatized central nervous system: hypoxia. Neurochem Res 2002;27:97-106.
- Dawson MR, Levine JM, Reynolds R. NG2-expressing cells in the central nervous system: are they oligodendroglial progenitors?. J Neurosci Res 2000;61:471-479.
- Emgard M, Hallin U, Karlsson J, Bahr BA, Brundin P, Blomgren K. Both apoptosis and necrosis occur early after intracerebral grafting of ventral mesencephalic tissue: a role for protease activation. J Neurochem 2003;86:1223-1232.
- Flax JD, Aurora S, Yang C, Simonin C, Wills AM, Billinghurst LL, Jendoubi M, Sidman RL, Wolfe JH, Kim SU, Snyder EY. Engraftable human neural stem cells respond to developmental cues, replace neurons, and express foreign genes. Nat Biotechnol 1998;16:1033-1039.
- Gahwiler BH, Capogna M, Debanne D, McKinney RA, Thompson SM. Organotypic slice cultures: a technique has come of age. Trends Neurosci 1997;20:471-477.
- Horner PJ, Power AE, Kempermann G, Kuhn HG, Palmer TD, Winkler J, Thal LJ, Gage FH. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J Neurosci 2000;20:2218-2228.
- Hwang DH, Kim BG, Kim EJ, Lee SI, Joo IS, Suh-Kim H, Sohn S, Kim SU. Transplantation of human neural stem cells transduced with Olig2 transcription factor improves locomotor recovery and enhances myelination in the white matter of rat spinal cord following contusive injury. BMC Neurosci 2009;10:117.
- Ishii K, Nakamura M, Dai H, Finn TP, Okano H, Toyama Y, Bregman BS. Neutralization of ciliary neurotrophic factor reduces astrocyte production from transplanted neural stem cells and promotes regeneration of corticospinal tract fibers in spinal cord injury. J Neurosci Res 2006;84:1669-1681.
- Jeong SW, Chu K, Jung KH, Kim SU, Kim M, Roh JK. Human Neural Stem Cell Transplantation Promotes Functional Recovery in Rats With Experimental Intracerebral Hemorrhage. Stroke 2003;34:2258-2263.
- Kim BG, Hwang DH, Lee SI, Kim EJ, Kim SU. Stem cell-based cell therapy for spinal cord injury. Cell Transplant 2007;16:357-366.
- Kim SU. Human neural stem cells genetically modified for brain repair in neurological disorders. Neuropathology 2004;24:159-171.
- Lee HJ, Kim KS, Kim EJ, Choi HB, Lee KH, Park IH, Ko Y, Jeong SW, Kim SU. Brain transplantation of immortalized human neural stem cells promotes functional recovery in mouse intracerebral hemorrhage stroke model. Stem Cells 2007;25:1204-1212.
- Lee SI, Kim BG, Hwang DH, Kim HM, Kim SU. Overexpression of Bcl-XL in human neural stem cells promotes graft survival and functional recovery following transplantation in spinal cord injury. J Neurosci Res 2009;87:3186-3197.
- McDonald JW, Belegu V. Demyelination and remyelination after spinal cord injury. J Neurotrauma 2006;23:345-359.
- McTigue DM, Wei P, Stokes BT. Proliferation of NG2-positive cells and altered oligodendrocyte numbers in the contused rat spinal cord. J Neurosci 2001;21:3392-3400.
- Meletis K, Barnabe-Heider F, Carlen M, Evergren E, Tomilin N, Shupliakov O, Frisen J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol 2008;6:e182.
- Meng XL, Shen JS, Ohashi T, Maeda H, Kim SU, Eto Y. Brain transplantation of genetically engineered human neural stem cells globally corrects brain lesions in the mucopolysaccharidosis type VII mouse. J Neurosci Res 2003;74:266-277.
- Monje ML, Mizumatsu S, Fike JR, Palmer TD. Irradiation induces neural precursor-cell dysfunction. Nat Med 2002;8:955-962.
- Noraberg J, Poulsen FR, Blaabjerg M, Kristensen BW, Bonde C, Montero M, Meyer M, Gramsbergen JB, Zimmer J. Organotypic hippocampal slice cultures for studies of brain damage, neuroprotection and neurorepair. Curr Drug Targets CNS Neurol Disord 2005;4:435-452.
- Okada S, Ishii K, Yamane J, Iwanami A, Ikegami T, Katoh H, Iwamoto Y, Nakamura M, Miyoshi H, Okano HJ, Contag CH, Toyama Y, Okano H.
In vivo imaging of engrafted neural stem cells: its application in evaluating the optimal timing of transplantation for spinal cord injury. FASEB 2005;19:1839-1841. - Okano H, Ogawa Y, Nakamura M, Kaneko S, Iwanami A, Toyama Y. Transplantation of neural stem cells into the spinal cord after injury. Semin Cell Dev Biol 2003;14:191-198.
- Pena F. Organotypic cultures as tool to test long-term effects of chemicals on the nervous system. Curr Med Chem 2010;17:987-1001.
- Rice T, Larsen J, Rivest S, Yong VW. Characterization of the early neuroinflammation after spinal cord injury in mice. J Neuropathol Exp Neurol 2007;66:184-195.
- Rossignol S, Schwab M, Schwartz M, Fehlings MG. Spinal cord injury: time to move?. J Neurosci 2007;27:11782-11792.
- Shihabuddin LS, Horner PJ, Ray J, Gage FH. Adult Spinal Cord Stem Cells Generate Neurons after Transplantation in the Adult Dentate Gyrus. J Neurosci 2000;20:8727-8735.
- Snyder EY, Park KI. Limitations in brain repair. Nat Med 2002;8:928-930.
- Stoppini L, Buchs PA, Muller D. A simple method for organotypic cultures of nervous tissue. J Neurosci Methods 1991;37:173-182.
- Tetzlaff W, Okon EB, Karimi-Abdolrezaee S, Hill CE, Sparling JS, Plemel JR, Plunet WT, Tsai EC, Baptiste D, Smithson LJ, Kawaja MD, Fehlings MG, Kwon BK. A Systematic Review of Cellular Transplantation Therapies for Spinal Cord Injury. J Neurotrauma 2010;27:1-72.
- Wang JK, Portbury S, Thomas MB, Barney S, Ricca DJ, Morris DL, Warner DS, Lo DC. Cardiac glycosides provide neuroprotection against ischemic stroke: discovery by a brain slice-based compound screening platform. Proc Natl Acad Sci U S A 2006;103:10461-10466.
- Yang H, Lu P, McKay HM, Bernot T, Keirstead H, Steward O, Gage FH, Edgerton VR, Tuszynski MH. Endogenous neurogenesis replaces oligodendrocytes and astrocytes after primate spinal cord injury. J Neurosci 2006;26:2157-2166.
- Yasuhara T, Matsukawa N, Hara K, Yu G, Xu L, Maki M, Kim S, Borlongan C. Transplantation of Human Neural Stem Cells Exerts Neuroprotection in a Rat Model of Parkinson's Disease. J Neurosci 2006;26:12497-12511.