Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Review Article
Exp Neurobiol 2013; 22(3): 149-157
Published online September 30, 2013
https://doi.org/10.5607/en.2013.22.3.149
© The Korean Society for Brain and Neural Sciences
Physiological and Pathological Significance of Dynamin-Related Protein 1 (Drp1)-Dependent Mitochondrial Fission in the Nervous System
Bongki Cho, So Yoen Choi, Hyo Min Cho, Hyun Jung Kim and Woong Sun*
Department of Anatomy, Korea University College of Medicine, Seoul 136-705, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-920-6404, FAX: 82-2-929-5696
e-mail: woongsun@korea.ac.kr
Abstract
Mitochondria are essential for proper neuronal morphogenesis and functions, as they are the major source of energy for neural development. The dynamic morphology of mitochondria determines the key functions of mitochondria. Several regulatory proteins such as dynamin-related protein 1 (Drp1) are required to maintain mitochondrial morphology via a balance between continuous fusion and fission. Activity of Drp1, a key regulator in mitochondrial fission, is modulated by multiple post-translation modifications and receptor interactions. In addition, numerous researches have revealed that the regulation of Drp1 activity and mitochondrial dynamics is closely associated with several neurodegenerative diseases such as Alzheimer's and Parkinson's diseases. In this article, we concisely review the recent findings about the biological importance of Drp1-mediated mitochondrial fission in neurons under physiological and pathological conditions.
Keywords: Drp1, neurodegeneration, mitochondria, fission, neuron
INTRODUCTION
Mitochondria support multiple aspects of neuronal development and functions by providing cellular energy, maintaining the intracellular calcium levels, generating reactive oxygen species and regulating the release of apoptogenic proteins such as cytochrome c and apoptosis-inducing factor [1]. Neurons are highly polarized cells with distinct axons and dendrites, and they have a high demand for energy to maintain neural circuits. Therefore, the quantity, quality and localization of mitochondria are all important for appropriate function of neurons [2]. For example, neuronal mitochondria are enriched in the synaptic structures and nodes of Ranvier where immediate supply of cellular energy and active buffering of calcium are required [3].
Recently, it has been demonstrated that morphological dynamics of mitochondria is also involved in the maintaining their function and distribution in neurons [1]. Mitochondrial morphology is determined by a balance between continuous fusion and fission [4], which is regulated by large GTPase dynamin-related proteins including Mitofusin 1/2 (Mfn1/2), Optic atrophy 1 (Opa1) and Dynamin-related protein 1 (Drp1) [5]. Mfn1/2 is located on the outer membrane of mitochondria (OMM), and binding and homo- or hetero-dimerization of mitofusins promote fusion of two neighboring OMM. Sequentially, Opa1, resided on the mitochondrial inner-membrane (IMM), fuses IMM by GTP hydrolysis. On the other hand, Drp1 is primarily localized at cytosol. For mitochondrial fission, Drp1 translocates from cytosol to OMM, and divides a mitochondrion into two pieces by GTP hydrolysis. Mitochondrial fission is predominantly controlled by the activity of Drp1, and the frequency of mitochondrial fission determines the quality as well as morphology of mitochondria. Because mitochondria are essential for the neuronal functions, increasing amounts of evidence support the idea that Drp1 plays critical roles in the physiological function and pathological progression of the nervous system.
MOLECULAR REGULATORY MECHANISMS FOR THE DRP1 FUNCTION
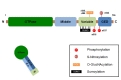
Drp1 protein has four domains: GTPase domain, middle domain variable domain, and GED domain (Fig. 1) [6]. Unlike other dynamin family proteins, however, Drp1 does not have a lipid-interacting pleckstrin homology domain [7], and thus the anchorage of Drp1 to the mitochondrial membrane is mediated by its receptor binding. Crystal structural study revealed that the variable domain of Drp1 acts as a hinge, forming a T-shaped dimer or tetramer [8, 9]. Therefore, fission-promoting activity of Drp1 is controlled by many post-translational modifications around the variable domain, including phosphorylation, sumoylation, ubiquitination and S-nitrosylation (Fig. 1, Table 1). Post-translational modifications and the alterations in its receptor functions also contribute to the activity of Drp1. S-nitrosylation is irreversible protein modification process, which is associated with neurodegenerative changes of proteins [10]. On the other hand, other post-translational modifications can be reversible, suggesting that these changes occur for the physiological control of mitochondrial dynamics.
Serine 637 (S637) of human Drp1 can be phosphorylated by protein kinase A (PKA), and it suppresses mitochondrial translocation and GTPase activity [11, 12]. On the other hand, Drp1S637 is dephosphorylated by calcineurin (PP2B), which promotes mitochondrial fragmentation [13]. In neurons, Drp1S637 can also be phosphorylated by CaMKIα following neuronal activation [14]. Interestingly, phosphorylation of Drp1 by CaMKIα conversely enhances the mitochondrial fragmentation, since phosphorylated Drp1 by CaMK1α shows higher affinity with mitochondrial effector molecule, Fis1. In addition, Rho-associated coiled-coil containing protein kinase 1 (ROCK1) also phosphorylates Drp1S637, resulting in mitochondrial fragmentation under hyperglycemic condition [15]. Considering that phosphorylation at the same site by different kinases showed entirely opposite effects on Drp1 activity, it appears that not only the status of Drp1 phosphorylation but also the type of upstream kinases determines the functional consequences. The precise mechanisms underlying these interesting phenomena remain to be explored.
Another phosphorylation site, serine 616 (S616), can also be phosphorylated by many kinases including CDK1 [16], ERK1/2 [17] and PKCδ [18]. Phosphorylation at this site enhances the activity of Drp1 under certain circumstances such as mitosis, high glucose or oxidative stress conditions, which results in mitochondrial fragmentation. However, when its phosphomimetic or phosphodefective mutant, Drp1(S616D) or Drp1(S616A) respectively, was simply overexpressed in HeLa cells, mutant Drp1 failed to alter mitochondrial morphology [13]. It implies that the effect of Drp1S616 phosphorylation may depend on intracellular context and/or upstream kinases rather than phosphorylation status. More recently, it has been identified that CDK5 can also phosphorylate Drp1S616, which induces mitochondrial fragmentation by the mobilization of Drp1 to mitochondria from microtubule [19]. However, CDK-dependent phosphorylation of Drp1S616 inhibits the oligomerization of Drp1 bound on microtubule or mitochondria. These results imply that CDK-dependent phosphorylation of Drp1S616 may has reciprocal effect on mitochondrial translocation and oligomerization of Drp1. Considering that neurons have high activity of CDK5, it is interesting to explore whether phosphorylation of Drp1S616 can serve as an activating or inactivating mechanism for Drp1 in the nervous system.
SUMOylation has been known to increase the stability of Drp1 on OMM that is required for efficient fission of mitochondria [20, 21]. For example, ectopic expression of SUMO1 stabilizes mitochondrial-targeted Drp1 in a Bax/Bak-dependent manner leading to mitochondrial fragmentation during apoptotic cell death [21]. Currently, the SUMOylation of Drp1 is regulated by SUMO E3 ligase MAPL [22] and SUMO protease SENP5 [23].
Drp1 activity can be regulated by ubiquitination, which is mediated by mitochondria-associated RING-finger E3 ubiquitin ligase (MARCH5/MITOL). Both overexpression of ligase activity-defective MARCH5 and gene knockdown induce hyperaccumulation of Drp1 on OMM, but leads to opposite effect which is mitochondrial elongation [24] or fragmentation [25]. These contradictory outputs result from diverse effects of MARCH5 on dynamin-related proteins; MARCH5 promotes Mfn2-mediated mitochondrial fusion, while it also ubiquitinates Drp1 [26]. In addition, Drp1 can also be ubiquitinated by Parkin implicated in Parkinson's disease [27]. This modification promotes proteasome-dependent degradation of Drp1, and pathogenic mutation or knockdown of Parkin induces increased level of Drp1 and aberrant mitochondrial fragmentation.
In rat neonatal cardiac myocytes, Drp1 can be O-GlcNAcylated by O-GlcNac-transferase at Threonine 585 and 586 in the variable domain [28]. This modification decreases the level of phosphorylated Drp1S637, which is consistent with a reciprocal relationship between O-GlcNAcylation and phosphorylation in several cases [29]. In fact, O-GlcNAcylation of Drp1 increases the levels of GTP-bound active Drp1 and leads to fragmentation of mitochondria.
MITOCHONDRIAL RECEPTORS FOR DRP1
Upon exposure to stimuli for mitochondrial fission, Drp1 is recruited from cytosol to putative fission sites on OMM. Recruitment of Drp1 is accompanied by several receptors on the OMM, and the interaction promotes Drp1-mediated mitochondrial fission. Fis1 was first identified as a Drp1 receptor in yeast [30]. In yeast, Fis1 is indirectly associated with Drp1 by two adaptor proteins, Mdv1 [31] and Caf4 [32]. However, mammalian homologues of Mdv1 and Caf4 have not been identified. In mammal, Drp1 interacts with Fis1 for mitochondrial fission [33], but Drp1 can also be recruited to mitochondria in a Fis1-independent manner [34, 35]. This implies alternative pathways of Drp1-mediated mitochondrial fission. Recent reports have demonstrated that mitochondrial fission factor (Mff) directly interacts with Drp1. In both
In addition, cytoskeleton is also involved in mitochondrial translocation of Drp1. Disruption of F-actin inhibits mitochondrial translocation of Drp1 under mitochondrial depolarization [39], and impaired microtubule sequestrates Drp1 away from mitochondria [19]. Interestingly, Drp1 also interacts with Mfn2 to inhibit mitochondrial fusion [40], suggesting that mitochondrial recruitment of Drp1 is involved in the mitochondria fusion as well. After mitochondrial recruitment, Drp1 forms ring- or spiral-like oligomers and weakly constricts at a putative fission site [41]. Finally, GTP hydrolysis promotes a complete fission of mitochondria [42].
MITOCHONDRIAL DYNAMICS AND NEURONAL FUNCTION UNDER PHYSIOLOGICAL CONDITIONS
Drp1 is a key player for the regulation of mitochondrial dynamics in neurons. Drp1 is highly expressed in post-mitotic neurons. RNA
Drp1 is essential for neuronal development, function and survival. Neuron-specific Drp1-/- mice died shortly after birth, resulting from developmental defects in nervous system [44]. In addition, a patient with dominant-negative mutation in Drp1 showed similar defects in brain development [48, 49]. These findings imply that Drp1-meditaed mitochondrial fission is required for neuronal development. During the development of nervous system, the length of mitochondria changes dynamically. Recently, we found that mitochondria in developing chick motoneurons progressively shortened
MITOCHONDRIAL DYNAMICS AND NEURONAL CELL DEATH AND PATHOLOGIES
The molecular links between Drp1 and cell death are first suggested by the fact that Drp1-containing fission foci also contains apoptosis-promoting Bcl-2 family molecules such as Bak, Bax and truncated Bid [21, 53-55]. In fact, mitochondrial fission is necessary for the execution of apoptosis. For example, suppression of Drp1 activity reduces mitochondrial fission and inhibits mitochondrial translocation of Bax which is essential for the execution of apoptosis [53, 56]. Conversely, the priming of mitochondrial fragmentation by Drp1 overexpression augments the extent of apoptosis [55], but Drp1 overexpression itself does not induce cell death in many cell types. Recently, we provided
Changes in the mitochondrial dynamics are found in many neurodegenerative diseases, including Alzheimer's disease (AD), Parkinson's disease (PD), Huntington's disease (HD) and amyotrophic lateral sclerosis (ALS). It has been postulated that the imbalance of mitochondrial fusion/fission is associated with disease-related mitochondrial dysfunctions [57]. In case of AD, mitochondrial fragmentation is progressively increased during the progression of disease in patients and transgenic mouse models [58, 59]. Upon induction of mitochondrial fragmentation, interactions between Drp1 and Aβ or phosphorylated Tau are also progressively increased [60, 61]. Although it is yet unclear how pathological changes enhance the interactions between Drp1 and AD-related proteins, it has been reported that post-translational modifications of Drp1 such as S-nitrosylation and phosphorylations are observed in AD patients [10, 62]. Therefore, it is plausible that AD-dependent modifications of Drp1 can enhance the fission activity as well as protein interactions.
In the pathology of PD, mitochondrial changes are well-documented. Especially, genetic mutations in the autophagic processes, such as Parkin and PINK, are frequently associated with PD. For example, Parkin and PINK play roles in the tagging of impaired mitochondria for autophagic degradation. Considering that mitochondrial fission is necessary for the efficient autophagic removal of damaged mitochondria, it has been proposed that mitochondrial fission machineries are affected by PD pathogenesis. As described in above, Parkin ubiquitinates Drp1 and promotes proteasome-dependent degradation of Drp1. Therefore, pathogenic mutation of Parkin results in the abnormal accumulation of Drp1, which promotes excessive mitochondrial fragmentation [27]. These mechanisms may be associated with PD-dependent mitochondrial dysfunctions. Drp1 also interacts with another PD-related molecule, leucine-rich repeat kinase 2 (LRRK2) [63]. This interaction enhances mitochondrial translocation of Drp1 and leads to excessive mitochondrial fragmentation.
The expression of Drp1 is increased in the striatum and cortex of HD patients, which may influence mitochondrial dysfunction in HD [64, 65]. Mutant huntingtin (mtHtt) abnormally recruits Drp1 on OMM and subsequently promotes GTPase activity of Drp1, resulting in excessive mitochondrial fragmentation [66, 67]. Recent report has revealed that hyper-activation of Drp1 is mediated by S-nitrosylation, as a result of nitric oxide produced by mtHtt [68], as also found in AD.
While mitochondrial fragmentation is not yet reported in ALS patients, an animal model of ALS, mutant SOD1 G93A (mSOD1) mouse, exhibited reduced mitochondrial length in motoneuron at the pre-symptomatic period [69, 70]. Furthermore, prevention of Drp1 activity significantly decreased cell death of spinal motoneuron in rats [71]. During the pathological process of ALS, the axonal transport of mitochondria is also impaired in affected motoneuron [72, 73]. These impairments are rescued by suppressing Drp1 [71], supporting the idea that mitochondrial fission machineries are tightly linked with axonal transport mechanism [3, 71, 74].
CONCLUSION
Dysregulation of Drp1-dependent mitochondrial fission is commonly observed in neurodegenerative diseases, despite the fact that no genetic mutation of Drp1 has been identified. Drp1 is therefore considered as a candidate target for a disease-modifying drug to enhance neuronal survival and their functions. For instance, chemical inhibitor of Drp1 (mdivi-1) can effectively suppressed retinal neurodegeneration following acute ischemia [75]. Suppression of basal Drp1 activity, however, causes impairments in synaptic plasticity and axonal integrity. Therefore, it is necessary to study the disease-specific modification of Drp1 and its implications, as it may provide a better insight to understand and treat neurodegenerative diseases.
Figures
{kind=link}
{kind=link}
Tables
{kind=link}
References
- Knott AB, Perkins G, Schwarzenbacher R, Bossy-Wetzel E. Mitochondrial fragmentation in neurodegeneration. Nat Rev Neurosci 2008;9:505-518.

- Liu L, Feng D, Chen G, Chen M, Zheng Q, Song P, Ma Q, Zhu C, Wang R, Qi W, Huang L, Xue P, Li B, Wang X, Jin H, Wang J, Yang F, Liu P, Zhu Y, Sui S, Chen Q. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat Cell Biol 2012;14:177-185.
- Baloh RH. Mitochondrial dynamics and peripheral neuropathy. Neuroscientist 2008;14:12-18.
- Chen H, Chan DC. Mitochondrial dynamics in mammals. Curr Top Dev Biol 2004;59:119-144.
- Praefcke GJ, McMahon HT. The dynamin superfamily: universal membrane tubulation and fission molecules?. Nat Rev Mol Cell Biol 2004;5:133-147.
- Okamoto K, Shaw JM. Mitochondrial morphology and dynamics in yeast and multicellular eukaryotes. Annu Rev Genet 2005;39:503-536.
- van der Bliek AM. Functional diversity in the dynamin family. Trends Cell Biol 1999;9:96-102.
- Ford MG, Jenni S, Nunnari J. The crystal structure of dynamin. Nature 2011;477:561-566.
- Fröhlich C, Grabiger S, Schwefel D, Faelber K, Rosenbaum E, Mears J, Rocks O, Daumke O. Structural insights into oligomerization and mitochondrial remodelling of dynamin 1-like protein. EMBO J 2013;32:1280-1292.
- Cho DH, Nakamura T, Fang J, Cieplak P, Godzik A, Gu Z, Lipton SA. S-nitrosylation of Drp1 mediates beta-amyloid-related mitochondrial fission and neuronal injury. Science 2009;324:102-105.
- Cribbs JT, Strack S. Reversible phosphorylation of Drp1 by cyclic AMP-dependent protein kinase and calcineurin regulates mitochondrial fission and cell death. EMBO Rep 2007;8:939-944.
- Chang CR, Blackstone C. Cyclic AMP-dependent protein kinase phosphorylation of Drp1 regulates its GTPase activity and mitochondrial morphology. J Biol Chem 2007;282:21583-21587.
- Cereghetti GM, Stangherlin A, Martins de Brito O, Chang CR, Blackstone C, Bernardi P, Scorrano L. Dephosphorylation by calcineurin regulates translocation of Drp1 to mitochondria. Proc Natl Acad Sci U S A 2008;105:15803-15808.
- Han XJ, Lu YF, Li SA, Kaitsuka T, Sato Y, Tomizawa K, Nairn AC, Takei K, Matsui H, Matsushita M. CaM kinase I alpha-induced phosphorylation of Drp1 regulates mitochondrial morphology. J Cell Biol 2008;182:573-585.
- Wang W, Wang Y, Long J, Wang J, Haudek SB, Overbeek P, Chang BH, Schumacker PT, Danesh FR. Mitochondrial fission triggered by hyperglycemia is mediated by ROCK1 activation in podocytes and endothelial cells. Cell Metab 2012;15:186-200.
- Taguchi N, Ishihara N, Jofuku A, Oka T, Mihara K. Mitotic phosphorylation of dynamin-related GTPase Drp1 participates in mitochondrial fission. J Biol Chem 2007;282:11521-11529.
- Yu T, Jhun BS, Yoon Y. High-glucose stimulation increases reactive oxygen species production through the calcium and mitogen-activated protein kinase-mediated activation of mitochondrial fission. Antioxid Redox Signal 2011;14:425-437.
- Qi X, Disatnik MH, Shen N, Sobel RA, Mochly-Rosen D. Aberrant mitochondrial fission in neurons induced by protein kinase C{delta} under oxidative stress conditions in vivo. Mol Biol Cell 2011;22:256-265.
- Strack S, Wilson TJ, Cribbs JT. Cyclin-dependent kinases regulate splice-specific targeting of dynamin-related protein 1 to microtubules. J Cell Biol 2013;201:1037-1051.
- Harder Z, Zunino R, McBride H. Sumo1 conjugates mitochondrial substrates and participates in mitochondrial fission. Curr Biol 2004;14:340-345.
- Wasiak S, Zunino R, McBride HM. Bax/Bak promote sumoylation of DRP1 and its stable association with mitochondria during apoptotic cell death. J Cell Biol 2007;177:439-450.
- Braschi E, Zunino R, McBride HM. MAPL is a new mitochondrial SUMO E3 ligase that regulates mitochondrial fission. EMBO Rep 2009;10:748-754.
- Zunino R, Schauss A, Rippstein P, Andrade-Navarro M, McBride HM. The SUMO protease SENP5 is required to maintain mitochondrial morphology and function. J Cell Sci 2007;120:1178-1188.
- Karbowski M, Neutzner A, Youle RJ. The mitochondrial E3 ubiquitin ligase MARCH5 is required for Drp1 dependent mitochondrial division. J Cell Biol 2007;178:71-84.
- Yonashiro R, Ishido S, Kyo S, Fukuda T, Goto E, Matsuki Y, Ohmura-Hoshino M, Sada K, Hotta H, Yamamura H, Inatome R, Yanagi S. A novel mitochondrial ubiquitin ligase plays a critical role in mitochondrial dynamics. EMBO J 2006;25:3618-3626.
- Nakamura N, Kimura Y, Tokuda M, Honda S, Hirose S. MARCH-V is a novel mitofusin 2- and Drp1-binding protein able to change mitochondrial morphology. EMBO Rep 2006;7:1019-1022.
- Wang H, Song P, Du L, Tian W, Yue W, Liu M, Li D, Wang B, Zhu Y, Cao C, Zhou J, Chen Q. Parkin ubiquitinates Drp1 for proteasome-dependent degradation: implication of dysregulated mitochondrial dynamics in Parkinson disease. J Biol Chem 2011;286:11649-11658.
- Gawlowski T, Suarez J, Scott B, Torres-Gonzalez M, Wang H, Schwappacher R, Han X, Yates JR, Hoshijima M, Dillmann W. Modulation of dynamin-related protein 1 (DRP1) function by increased O-linked-beta-N-acetylglucosamine modification (O-GlcNAc) in cardiac myocytes. J Biol Chem 2012;287:30024-30034.
- Butkinaree C, Park K, Hart GW. O-linked beta-N-acetylglucosamine (O-GlcNAc): Extensive crosstalk with phosphorylation to regulate signaling and transcription in response to nutrients and stress. Biochim Biophys Acta 2010;1800:96-106.
- Mozdy AD, McCaffery JM, Shaw JM. Dnm1p GTPase-mediated mitochondrial fission is a multi-step process requiring the novel integral membrane component Fis1p. J Cell Biol 2000;151:367-380.
- Tieu Q, Okreglak V, Naylor K, Nunnari J. The WD repeat protein, Mdv1p, functions as a molecular adaptor by interacting with Dnm1p and Fis1p during mitochondrial fission. J Cell Biol 2002;158:445-452.
- Griffin EE, Graumann J, Chan DC. The WD40 protein Caf4p is a component of the mitochondrial fission machinery and recruits Dnm1p to mitochondria. J Cell Biol 2005;170:237-248.
- Yoon Y, Krueger EW, Oswald BJ, McNiven MA. The mitochondrial protein hFis1 regulates mitochondrial fission in mammalian cells through an interaction with the dynamin-like protein DLP1. Mol Cell Biol 2003;23:5409-5420.
- Lee S, Jeong SY, Lim WC, Kim S, Park YY, Sun X, Youle RJ, Cho H. Mitochondrial fission and fusion mediators, hFis1 and OPA1, modulate cellular senescence. J Biol Chem 2007;282:22977-22983.
- Wasiak S, Zunino R, McBride HM. Bax/Bak promote sumoylation of DRP1 and its stable association with mitochondria during apoptotic cell death. J Cell Biol 2007;177:439-450.
- Otera H, Wang C, Cleland MM, Setoguchi K, Yokota S, Youle RJ, Mihara K. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J Cell Biol 2010;191:1141-1158.
- Palmer CS, Osellame LD, Laine D, Koutsopoulos OS, Frazier AE, Ryan MT. MiD49 and MiD51, new components of the mitochondrial fission machinery. EMBO Rep 2011;12:565-573.
- Zhao J, Liu T, Jin S, Wang X, Qu M, Uhlén P, Tomilin N, Shupliakov O, Lendahl U, Nistér M. Human MIEF1 recruits Drp1 to mitochondrial outer membranes and promotes mitochondrial fusion rather than fission. EMBO J 2011;30:2762-2778.
- De Vos KJ, Allan VJ, Grierson AJ, Sheetz MP. Mitochondrial function and actin regulate dynamin-related protein 1-dependent mitochondrial fission. Curr Biol 2005;15:678-683.
- Huang P, Galloway CA, Yoon Y. Control of mitochondrial morphology through differential interactions of mitochondrial fusion and fission proteins. PLoS One 2011;6:e20655.
- Ingerman E, Perkins EM, Marino M, Mears JA, McCaffery JM, Hinshaw JE, Nunnari J. Dnm1 forms spirals that are structurally tailored to fit mitochondria. J Cell Biol 2005;170:1021-1027.
- Mears JA, Lackner LL, Fang S, Ingerman E, Nunnari J, Hinshaw JE. Conformational changes in Dnm1 support a contractile mechanism for mitochondrial fission. Nat Struct Mol Biol 2011;18:20-26.
- Li Z, Okamoto K, Hayashi Y, Sheng M. The importance of dendritic mitochondria in the morphogenesis and plasticity of spines and synapses. Cell 2004;119:873-887.
- Ishihara N, Nomura M, Jofuku A, Kato H, Suzuki SO, Masuda K, Otera H, Nakanishi Y, Nonaka I, Goto Y, Taguchi N, Morinaga H, Maeda M, Takayanagi R, Yokota S, Mihara K. Mitochondrial fission factor Drp1 is essential for embryonic development and synapse formation in mice. Nat Cell Biol 2009;11:958-966.
- Choi SY, Kim JY, Kim HW, Cho B, Cho HM, Oppenheim RW, Kim H, Rhyu IJ, Sun W. Drp1-mediated mitochondrial dynamics and survival of developing chick motoneurons during the period of normal programmed cell death. FASEB J 2013;27:51-62.
- Dickey AS, Strack S. PKA/AKAP1 and PP2A/Bbeta2 regulate neuronal morphogenesis via Drp1 phosphorylation and mitochondrial bioenergetics. J Neurosci 2011;31:15716-15726.
- Li H, Chen Y, Jones AF, Sanger RH, Collis LP, Flannery R, McNay EC, Yu T, Schwarzenbacher R, Bossy B, Bossy-Wetzel E, Bennett MV, Pypaert M, Hickman JA, Smith PJ, Hardwick JM, Jonas EA. Bcl-xL induces Drp1-dependent synapse formation in cultured hippocampal neurons. Proc Natl Acad Sci U S A 2008;105:2169-2174.
- Waterham HR, Koster J, van Roermund CW, Mooyer PA, Wanders RJ, Leonard JV. A lethal defect of mitochondrial and peroxisomal fission. N Engl J Med 2007;356:1736-1741.
- Wakabayashi J, Zhang Z, Wakabayashi N, Tamura Y, Fukaya M, Kensler TW, Iijima M, Sesaki H. The dynamin-related GTPase Drp1 is required for embryonic and brain development in mice. J Cell Biol 2009;186:805-816.
- Cheng A, Wan R, Yang JL, Kamimura N, Son TG, Ouyang X, Luo Y, Okun E, Mattson MP. Involvement of PGC-1alpha in the formation and maintenance of neuronal dendritic spines. Nat Commun 2012;3:1250.
- Chang DT, Reynolds IJ. Mitochondrial trafficking and morphology in healthy and injured neurons. Prog Neurobiol 2006;80:241-268.
- Chang DT, Reynolds IJ. Differences in mitochondrial movement and morphology in young and mature primary cortical neurons in culture. Neuroscience 2006;141:727-736.
- Montessuit S, Somasekharan SP, Terrones O, Lucken-Ardjomande S, Herzig S, Schwarzenbacher R, Manstein DJ, Bossy-Wetzel E, Basañez G, Meda P, Martinou JC. Membrane remodeling induced by the dynamin-related protein Drp1 stimulates Bax oligomerization. Cell 2010;142:889-901.
- Cassidy-Stone A, Chipuk JE, Ingerman E, Song C, Yoo C, Kuwana T, Kurth MJ, Shaw JT, Hinshaw JE, Green DR, Nunnari J. Chemical inhibition of the mitochondrial division dynamin reveals its role in Bax/Bak-dependent mitochondrial outer membrane permeabilization. Dev Cell 2008;14:193-204.
- Brooks C, Cho SG, Wang CY, Yang T, Dong Z. Fragmented mitochondria are sensitized to Bax insertion and activation during apoptosis. Am J Physiol Cell Physiol 2011;300:C447-C455.
- Karbowski M, Lee YJ, Gaume B, Jeong SY, Frank S, Nechushtan A, Santel A, Fuller M, Smith CL, Youle RJ. Spatial and temporal association of Bax with mitochondrial fission sites, Drp1, and Mfn2 during apoptosis. J Cell Biol 2002;159:931-938.
- Knott AB, Bossy-Wetzel E. Impairing the mitochondrial fission and fusion balance: a new mechanism of neurodegeneration. Ann N Y Acad Sci 2008;1147:283-292.
- Su B, Wang X, Bonda D, Perry G, Smith M, Zhu X. Abnormal mitochondrial dynamics--a novel therapeutic target for Alzheimer's disease?. Mol Neurobiol 2010;41:87-96.
- Wang X, Su B, Lee HG, Li X, Perry G, Smith MA, Zhu X. Impaired balance of mitochondrial fission and fusion in Alzheimer's disease. J Neurosci 2009;29:9090-9103.
- Manczak M, Calkins MJ, Reddy PH. Impaired mitochondrial dynamics and abnormal interaction of amyloid beta with mitochondrial protein Drp1 in neurons from patients with Alzheimer's disease: implications for neuronal damage. Hum Mol Genet 2011;20:2495-2509.
- Manczak M, Reddy PH. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimer's disease neurons: implications for mitochondrial dysfunction and neuronal damage. Hum Mol Genet 2012;21:2538-2547.
- Wang S, Song J, Tan M, Albers KM, Jia J. Mitochondrial fission proteins in peripheral blood lymphocytes are potential biomarkers for Alzheimer's disease. Eur J Neurol 2012;19:1015-1022.
- Wang X, Yan MH, Fujioka H, Liu J, Wilson-Delfosse A, Chen SG, Perry G, Casadesus G, Zhu X. LRRK2 regulates mitochondrial dynamics and function through direct interaction with DLP1. Hum Mol Genet 2012;21:1931-1944.
- Shirendeb U, Reddy AP, Manczak M, Calkins MJ, Mao P, Tagle DA, Reddy PH. Abnormal mitochondrial dynamics, mitochondrial loss and mutant huntingtin oligomers in Huntington's disease: implications for selective neuronal damage. Hum Mol Genet 2011;20:1438-1455.
- Costa V, Giacomello M, Hudec R, Lopreiato R, Ermak G, Lim D, Malorni W, Davies KJ, Carafoli E, Scorrano L. Mitochondrial fission and cristae disruption increase the response of cell models of Huntington's disease to apoptotic stimuli. EMBO Mol Med 2010;2:490-503.
- Song W, Chen J, Petrilli A, Liot G, Klinglmayr E, Zhou Y, Poquiz P, Tjong J, Pouladi MA, Hayden MR, Masliah E, Ellisman M, Rouiller I, Schwarzenbacher R, Bossy B, Perkins G, Bossy-Wetzel E. Mutant huntingtin binds the mitochondrial fission GTPase dynamin-related protein-1 and increases its enzymatic activity. Nat Med 2011;17:377-382.
- Shirendeb UP, Calkins MJ, Manczak M, Anekonda V, Dufour B, McBride JL, Mao P, Reddy PH. Mutant huntingtin's interaction with mitochondrial protein Drp1 impairs mitochondrial biogenesis and causes defective axonal transport and synaptic degeneration in Huntington's disease. Hum Mol Genet 2012;21:406-420.
- Haun F, Nakamura T, Shiu AD, Cho DH, Tsunemi T, Holland EA, La Spada AR, Lipton SA. S-nitrosylation of dynamin-related protein 1 mediates mutant huntingtin-induced mitochondrial fragmentation and neuronal injury in Huntington's disease. Antioxid Redox Signal 2013
- Wong PC, Pardo CA, Borchelt DR, Lee MK, Copeland NG, Jenkins NA, Sisodia SS, Cleveland DW, Price DL. An adverse property of a familial ALS-linked SOD1 mutation causes motor neuron disease characterized by vacuolar degeneration of mitochondria. Neuron 1995;14:1105-1116.
- Kong J, Xu Z. Massive mitochondrial degeneration in motor neurons triggers the onset of amyotrophic lateral sclerosis in mice expressing a mutant SOD1. J Neurosci 1998;18:3241-3250.
- Song W, Song Y, Kincaid B, Bossy B, Bossy-Wetzel E. Mutant SOD1G93A triggers mitochondrial fragmentation in spinal cord motor neurons: neuroprotection by SIRT3 and PGC-1alpha. Neurobiol Dis 2013;51:72-81.
- Millecamps S, Julien JP. Axonal transport deficits and neurodegenerative diseases. Nat Rev Neurosci 2013;14:161-176.
- Bilsland LG, Sahai E, Kelly G, Golding M, Greensmith L, Schiavo G. Deficits in axonal transport precede ALS symptoms in vivo. Proc Natl Acad Sci U S A 2010;107:20523-20528.
- Chen H, Chan DC. Mitochondrial dynamics--fusion, fission, movement, and mitophagy--in neurodegenerative diseases. Hum Mol Genet 2009;18:R169-R176.
- Park SW, Kim KY, Lindsey JD, Dai Y, Heo H, Nguyen DH, Ellisman MH, Weinreb RN, Ju WK. A selective inhibitor of drp1, mdivi-1, increases retinal ganglion cell survival in acute ischemic mouse retina. Invest Ophthalmol Vis Sci 2011;52:2837-2843.