Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2013; 22(4): 315-321
Published online December 30, 2013
https://doi.org/10.5607/en.2013.22.4.315
© The Korean Society for Brain and Neural Sciences
Zinc-Triggered Induction of Tissue Plasminogen Activator and Plasminogen in
Endothelial Cells and Pericytes
Mun-Kyung Cho, Eun-Sun Sun and Yang-Hee Kim*
Department of Molecular Biology, Sejong University, Seoul 143-747, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-3408-3648, FAX: 82-2-3408-4336
e-mail: yhkim@sejong.ac.kr
Cerebral amyloid angiopathy (CAA) is common in patients with Alzheimer's disease (AD) and may contribute to cerebral hemorrhage. We previously demonstrated that tissue plasminogen activator (tPA) and plasminogen (PLG) accumulated at the periphery of compact amyloid-cored plaques and in the walls of CAA-containing blood vessels in the brains of Tg2576 mice, a widely used AD mouse model. We had also observed that zinc-triggered tPA and PLG induction were observed in mouse cortical cultures. Because zinc also accumulates in amyloid plaques and blood vessel walls in AD brains, we examined whether zinc increases mRNA and protein levels of tPA and PLG in brain endothelial cells and pericytes. Four hours after the exposure of brain endothelial cells (bEnd.3) to 40 µM zinc, the mRNA and protein expressions of tPA and its substrate PLG were significantly increased. In the case of brain pericyte cultures, increases in tPA and PLG expression were also detected 2 hr after treatment. However, amyloid-β (Aβ)1-42 oligomers did not augment tPA and PLG expression in bEnd.3 cells and pericytes, suggesting that zinc but not Aβ induces tPA and PLG accumulation in CAA found in the AD brain.
Keywords: zinc, amyloid-β (Aβ), tissue plasminogen activator (tPA), plasminogen (PLG), cerebral amyloid angiopathy (CAA)
INTRODUCTION
Amyloid-β (Aβ) accumulates in the brain of Alzheimer's disease (AD) patients as parenchymal plaque deposits [1] and cerebral amyloid angiopathy (CAA) in blood vessel walls [2]. Aβ is a 39~43 amino acid peptide cleaved from amyloid β-protein precursor (AβPP), by a β-secretase and a γ-secretase [3-8]. Increased levels of the hydrophobic peptides Aβ1-40 or Aβ1-42 in AD brain results in their self-assembly into insoluble and β-sheet-containing fibrils, which are ultimately deposited in the brain parenchyma and vessel walls [1, 9].
Zinc plays a critical role in brain parenchymal and vascular Aβ aggregation in AD [10-12]. It is concentrated in amyloid plaques in AD, as well as in an AD transgenic mouse model (Tg2576)[13, 14]. The chelation of zinc resolubilizes Aβ from post-mortem AD tissue [15] and reduces brain amyloid burden in Tg2576 mice [16]. In addition, genetic deletion of zinc transporter 3 (ZnT3), which inserts exchangeable zinc into synaptic vesicles, markedly attenuated amyloid deposition in Tg2576 mice [17], suggesting a key role of synaptic zinc in Aβ plaque formation. Consistent with this, histochemically reactive zinc [detected by N-(6-methoxy-8-quinolyl)-p-carboxybenzoylsulphonamide (TFL-Zn)] was also detected in CAA in Tg2576 mice, and noticeable reduction of CAA was observed in ZnT3-depleted Tg2576 mice (
Tissue plasminogen activator (tPA) is a serine protease involved in thrombolysis [18-20]. tPA converts plasminogen (PLG) to plasmin, the most potent and reactive serine protease [18]. The tPA-PLG-plasmin cascade system is involved in various physiological and pathological processes, including thrombolysis, embryo development, wound healing, and cancer progression [18, 21-23]. In the central nervous system, the tPA/PLG/plasmin system exerts additional beneficial and detrimental functions. Whereas tPA induces neurite outgrowth and neuronal survival [24, 25], tPA exacerbates mild excitotoxicity by proteolytically up-regulating N-methyl-D aspartate receptor function [26]. Because of the potent protease action of the tPA/PLG/plasmin system, the expression levels of tPA and its substrate, PLG, are tightly regulated [18]. However, zinc neurotoxicity, a possible underlying neuronal death mechanism for brain ischemia and seizure [27, 28], was markedly attenuated by tPA [29]. In addition, the tPA/PLG/plasmin system is involved in long-term potentiation [30, 31] via cleavage of pro-brain-derived neurotrophic factor (BDNF) to mature BDNF [32], proteolytic degradation of laminin [33, 34], and lipoprotein receptor-mediated induction of matrix metalloproteinase (MMP) [35]. We previously reported that tPA/PLG co-accumulated at the periphery of amyloid deposits and in CAA-containing blood vessel walls in Tg2576 mice [36]. Although tPA/PLG/plasmin system degrades Aβ fibrils and reduces the overall Aβ burden [37, 38], it also increases the risk of hemorrhage. In mouse cortical cultures, we found that mRNA and protein levels of tPA and PLG were regulated by zinc via MMP activation and BDNF secretion [39]. However, it is not clear how tPA and PLG are upregulated in CAA. In the present study, we investigated whether zinc or Aβ that is co-accumulated into CAA with tPA and PLG play a key role in regulating tPA and PLG expression in endothelial cells and pericytes.
MATERIALS AND METHODS
Immortalized mouse brain endothelial bEnd.3 cells were purchased from the American Type Culture Collection (Manassas, VA, USA) and maintained in Dulbecco's Modified Eagle's Medium (DMEM; Gibco/Invitrogen, Carlsbad, CA USA) containing 10% FBS (Hyclone, Logan, UT, USA), and antibiotics (100 units/mL penicillin and 100 µg/mL streptomycin) at 37℃ in a humidified 5% CO2 atmosphere.
Mouse brain pericytes were obtained from 8-week old mouse cortical tissue [40]. Dissociated cortical tissues were digested with an enzymatic solution (15 units/ml papain [Worthington Biochemical Corp., Lakewood, NJ, USA], 2,000 units/ml DNase I [Worthington], 1.1 mM EDTA, 0.067 mM mercaptoethanol, and 5.5 mM cystein-HCl in Earle's Balanced Salt Solution [EBSS; Gibco/Invitrogen, USA]) in a 37℃ water bath for 30 min, and then homogenized by passage through an 18-gauge needle five times, followed by five passages through a 21-gauge needle. The homogenized cortical tissues were mixed with 1.7 volumes of Dulbecco's phosphate-buffered saline (DPBS) containing 22% bovine serum albumin (BSA) and then centrifuged at 4,000 rpm for 10 min. The cell pellet was re-suspended in endothelial growth medium (ECGM; endothelial basal medium supplemented with 2% FBS, hydrocortisone, human epidermal growth factor, vascular endothelial growth factor, human fibroblast growth factor-B, R3-insulin-like growth factor-1 [R3-IGF-1], ascorbic acid, heparin, and gentamicin/amphotericin [all from LONZA, Basel, Switzerland]) and then plated onto two wells of a 6-well plate that had been coated with 300 µg/cm2 collagen (Sigma, St. Louis, MO, USA). The cells were kept in ECGM during the first two passages, and then incubated in 2% FBS-pericyte medium (ScienCell Research Laboratories, Carlsbad, CA, USA) for the next 3~4 passages and then finally maintained in 10% FBS-DMEM at 37℃ in a humidified 5% CO2 atmosphere. Primary mouse brain pericytes that were between 10 and 30 passages were used for experiments. Before use, each culture was assessed for purity by performing immunocytochemistry with anti-CD31 (an endothelial cell marker) and anti-CD146 (a pericyte marker); we confirmed that 100% of the cells were pericytes, and endothelial cells were absent in these cultures (Fig. 1).
Mouse brain endothelial cells (bEnd.3) and pericytes were exposed to zinc (ZnCl2) or Aβ (Aβ1-42 [BACHEM, Bubendorf, Switzerland]) in serum-free MEM (Eagle's minimal essential medium, Gibco/Invitrogen). To prepare Aβ oligomers [41], 1 mg Aβ1-42 was suspended at an initial concentration of 1 mM in 222 µl hexafluoroisopropanol (HFIP; J.T. Baker, Phillipsburg, NJ, USA), using a Hamilton syringe and then incubated at room temperature (RT) for 2 hr. HFIP was evaporated off in a fume hood. The peptide was resuspended at a concentration of 5 mM by addition of 44 µl dimethyl sulfoxide (DMSO; ATCC). The peptide solution was vortexed and sonicated with a water ultrasound sonicator at RT for 10 min. For oligomerization, Aβ1-42 peptide was diluted in 440 µl EBSS at a concentration of 500 µM and then incubated at 4℃ for at least 24 h. Then, Aβ1-42 peptide solution was centrifuged at 11,000×g for 2 min, and the supernatant was collected for experimental use. Immediately before their use, oligomers were sonicated with a water ultrasound sonicator for 10 min at RT.
Cells were fixed in 4% paraformaldehyde at 4℃ for 30 min and permeabilized with 0.1% Triton X-100 in DPBS. After blocking with normal serum in DPBS, cultures were incubated with CD31 (endothelial cell marker; Abcam, Cambridge UK) or CD146 (pericytes marker; Abcam) antibodies at 4℃ overnight. Cultures were washed and incubated with an Alexa Fluor 488-conjugated secondary antibody for 2 hr. Cells were observed under a fluorescence microscope (EVOSfl, Advanced Microscopy Group, Bothell, WA, USA).
Total RNA was isolated with a TRIzol Reagent (Invitrogen) according to the manufacturer's manual and reverse-transcribed into cDNA using iScript™ cDNA Synthesis Kit (Bio-Rad, Hercules, CA USA). Reverse-transcriptase polymerase chain reaction (RT-PCR) was performed with the following primer sets specific for mouse sequences: tPA primer (forward, 5'-TGCCTTCCTCTTCCTCTTCTACAG-3'; reverse, 5'-AGATACCCCTTCCTTCTCGTGG-3'), PLG primer (forward, 5'-CAGCAAGACTTCCTCCATCATC-3'; reverse, 5'-CCGTCAGGGTTGCGGCAATAAT-3'), β-actin primer (forward, 5'-TCTACAAATGTGGCTGAGGAC-3'; reverse, 5'-CCTGGGCCATTCAGAAATTA-3'). Primer sequences were designed based on published cDNA sequences. PCR was carried out for 25 cycles (94℃ for 60 sec, 60℃ for 90 sec, and 72℃ for 60 sec) for tPA and 35 cycles (94℃ for 60 sec, 61℃ for 60 sec, and 72℃ for 60 sec) for PLG.
Cell lysates were prepared in lysis buffer (62.5 mM Tris [pH 6.8], 6 M urea, 2% sodium dodecyl sulfate [SDS], 5% mercaptoethanol, 20 mM dithiothreitol, 0.005% bromophenol blue, 10% glycerol) containing freshly prepared protease and phosphatase inhibitors (2 µg/mL aprotinin, 1 µg/mL leupeptin, 1 µg/ml pepstatin A, 1 mM phenylmethylsulfonyl fluoride, 5 mM NaF, 1 mM Na3VO4, and 10 mM Na4P2O7). For each sample, 30 µg total protein was separated by SDS-polyacrylamide gels (8%) under reducing conditions and immunoblotted with antibodies against mouse tPA (ab28374, Abcam), PLG (ab154560, Abcam), or actin (A5060, Sigma). Enhanced chemiluminescence reagents (Intron Biotechnology, Sangdaewon-dong, South Korea) were used to visualize the immunoreactive bands.
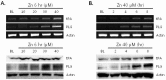
To determine whether zinc increases tPA and PLG expression in brain endothelial cells, we treated bEnd.3 cultures with sub-lethal doses of zinc (10~40 µM). Six hours later, increases in mRNA and protein levels of tPA and PLG were detected for zinc concentrations greater than 10 µM (Fig. 2A). To determine the time points of tPA and PLG induction by zinc, we examined tPA and PLG expression levels from 2 to 8 hr after bEnd.3 cells were treated with zinc. We observed substantial inductions of tPA and PLG beginning 4 hr after zinc exposure to bEnd.3 cultures (Fig. 2B).
Next, we examined whether zinc could also augment the expression levels of tPA and PLG in mouse brain pericytes. Sub-lethal dose of zinc (10~40 µM) also increased mRNA and protein levels of tPA and PLG in pericytes (Fig. 3A). We detected increased mRNA and protein levels of tPA 2 hr after pericytes were treated with zinc (Fig. 3B), which is earlier than observed for bEnd.3 cells (Fig. 2B). Distinct induction of PLG by zinc in pericytes was observed from 4 hr on (Fig. 3B).
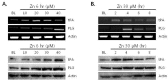
Because we previously reported co-accumulation of tPA, PLG, and zinc, as well as Aβ in amyloid plaques in the brains and blood vessel walls of Tg2576 AD model mice [36], we investigated whether tPA and PLG induction was triggered by Aβ in bEnd.3 or pericyte cultures. In contrast with zinc-induced expression of tPA and PLG, 10 µM Aβ treatment did not increase tPA or PLG expression levels in bEnd.3 or pericyte cultures (Fig. 4). These results indicate that induction of tPA and PLG in endothelial cells and pericytes are largely dependent on zinc and are not influenced by Aβ.
Ours results demonstrate that mRNA and protein levels of tPA and PLG were noticeably increased by zinc in both brain endothelial cells and pericytes. However, although Aβ co-accumulates with zinc, tPA, and PLG in amyloid deposits and CAA in Tg2576 mice, Aβ treatment did not affect tPA or PLG expression in bEnd.3 endothelial cultures or primary brain pericytes (Fig. 4). Although tPA's protease activity is stimulated by assembled Aβ in vitro [42], we found that tPA and PLG expression levels in bEnd.3 endothelial cells and pericytes were not affected by treatment with Aβ1-42 oligomers. This result suggested that the major function of tPA/PLG induction in bEnd.3 endothelial cells and pericytes may not be the degradation of Aβ. One of the possible roles of tPA induction could be the attenuation of zinc-induced damage, because zinc neurotoxicity was almost completely blocked by tPA [29, 43]. Emmetsberger and colleagues showed that reduction of zinc toxicity by tPA was mediated by promotion of zinc uptake into cells via ZIP4 [43]. Taken together, tPA/PLG system in bEnd.3 endothelial cells and pericytes may be largely related to zinc homeostasis rather than Aβ clearance.
However, it is still possible that tPA/PLG/plasmin cascade system may help clear Aβ. While hippocampal injections of Aβ into mice lacking either tPA or PLG persist and induce neuronal damage, Aβ injected into normal mice is cleared and does not cause neuronal damage [44]. tPA and PLG may also increase the risk of hemorrhage in CAA. Upregulation of plasminogen activator inhibitor-1 (PAI-1), an inducible protein inhibitor of tPA, contributes to the protective effects of tPA/PLG/plasmin system in wild-type mice. To understand whether zinc-induced tPA and PLG expression in endothelial cells and pericytes have beneficial or detrimental effects, zinc-triggered PAI-1 induction should be examined in future studies.
ZnT3 and synaptic zinc play key roles in brain parenchymal and vascular Aβ deposition; deletion of ZnT3 dramatically reduced amyloid plaques and CAA in Tg2576 AD model mice [12, 17]. To assess the relevance of zinc-triggered induction of tPA and PLG in vascular cell cultures to CAA in AD, tPA and PLG expression levels should be examined in ZnT3-disrupted Tg2576 mice (
We previously demonstrated that zinc-triggered tPA and PLG induction in mouse cortical cultures was mediated by MMP activation and a subsequent increase of pro-BDNF cleavage [39]. Upregulation of BDNF induces tropomyosin receptor kinase (Trk) signaling, which then increases tPA and PLG expression in cortical neuronal cultures [13]. Further studies are needed to understand which signaling pathways are involved in zinc-triggered tPA and PLG induction in endothelial cells and pericytes. Such research will provide important insights into AD research and elucidate the underlying mechanisms of CAA.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc Natl Acad Sci U S A 1985;82:4245-4249.

- Glenner GG, Wong CW. Alzheimer's disease: initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem Biophys Res Commun 1984;120:885-890.
- Kang J, Lemaire HG, Unterbeck A, Salbaum JM, Masters CL, Grzeschik KH, Multhaup G, Beyreuther K, Müller-Hill B. The precursor of Alzheimer's disease amyloid A4 protein resembles a cell-surface receptor. Nature 1987;325:733-736.
- Vassar R, Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA, Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, Luo Y, Fisher S, Fuller J, Edenson S, Lile J, Jarosinski MA, Biere AL, Curran E, Burgess T, Louis JC, Collins F, Treanor J, Rogers G, Citron M. Beta-secretase cleavage of Alzheimer's amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999;286:735-741.
- Yan R, Bienkowski MJ, Shuck ME, Miao H, Tory MC, Pauley AM, Brashier JR, Stratman NC, Mathews WR, Buhl AE, Carter DB, Tomasselli AG, Parodi LA, Heinrikson RL, Gurney ME. Membrane-anchored aspartyl protease with Alzheimer's disease beta-secretase activity. Nature 1999;402:533-537.
- Sinha S, Anderson JP, Barbour R, Basi GS, Caccavello R, Davis D, Doan M, Dovey HF, Frigon N, Hong J, Jacobson-Croak K, Jewett N, Keim P, Knops J, Lieberburg I, Power M, Tan H, Tatsuno G, Tung J, Schenk D, Seubert P, Suomensaari SM, Wang S, Walker D, Zhao J, McConlogue L, John V. Purification and cloning of amyloid precursor protein beta-secretase from human brain. Nature 1999;402:537-540.
- De Strooper B, Saftig P, Craessaerts K, Vanderstichele H, Guhde G, Annaert W, Von Figura K, Van Leuven F. Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nature 1998;391:387-390.
- Wolfe MS, Xia W, Ostaszewski BL, Diehl TS, Kimberly WT, Selkoe DJ. Two transmembrane aspartates in presenilin-1 required for presenilin endoproteolysis and gamma-secretase activity. Nature 1999;398:513-517.
- Selkoe DJ. Alzheimer's disease: genes, proteins, and therapy. Physiol Rev 2001;81:741-766.
- Bush AI, Pettingell WH, Multhaup G, d Paradis M, Vonsattel JP, Gusella JF, Beyreuther K, Masters CL, Tanzi RE. Rapid induction of Alzheimer A beta amyloid formation by zinc. Science 1994;265:1464-1467.
- Huang X, Atwood CS, Moir RD, Hartshorn MA, Vonsattel JP, Tanzi RE, Bush AI. Zinc-induced Alzheimer's Abeta1-40 aggregation is mediated by conformational factors. J Biol Chem 1997;272:26464-26470.
- Friedlich AL, Lee JY, van Groen T, Cherny RA, Volitakis I, Cole TB, Palmiter RD, Koh JY, Bush AI. Neuronal zinc exchange with the blood vessel wall promotes cerebral amyloid angiopathy in an animal model of Alzheimer's disease. J Neurosci 2004;24:3453-3459.
- Lee JY, Mook-Jung I, Koh JY. Histochemically reactive zinc in plaques of the Swedish mutant beta-amyloid precursor protein transgenic mice. J Neurosci 1999;19:RC10.
- Suh SW, Jensen KB, Jensen MS, Silva DS, Kesslak PJ, Danscher G, Frederickson CJ. Histochemically-reactive zinc in amyloid plaques, angiopathy, and degenerating neurons of Alzheimer's diseased brains. Brain Res 2000;852:274-278.
- Cherny RA, Legg JT, McLean CA, Fairlie DP, Huang X, Atwood CS, Beyreuther K, Tanzi RE, Masters CL, Bush AI. Aqueous dissolution of Alzheimer's disease Abeta amyloid deposits by biometal depletion. J Biol Chem 1999;274:23223-23228.
- Cherny RA, Atwood CS, Xilinas ME, Gray DN, Jones WD, McLean CA, Barnham KJ, Volitakis I, Fraser FW, Kim Y, Huang X, Goldstein LE, Moir RD, Lim JT, Beyreuther K, Zheng H, Tanzi RE, Masters CL, Bush AI. Treatment with a copper-zinc chelator markedly and rapidly inhibits beta-amyloid accumulation in Alzheimer's disease transgenic mice. Neuron 2001;30:665-676.
- Lee JY, Cole TB, Palmiter RD, Suh SW, Koh JY. Contribution by synaptic zinc to the gender-disparate plaque formation in human Swedish mutant APP transgenic mice. Proc Natl Acad Sci U S A 2002;99:7705-7710.
- Kane KK. Fibrinolysis--a review. Ann Clin Lab Sci 1984;14:443-449.
- Danglot G, Vinson D, Chapeville F. Qualitative and quantitative distribution of plasminogen activators in organs from healthy adult mice. FEBS Lett 1986;194:96-100.
- Sappino AP, Madani R, Huarte J, Belin D, Kiss JZ, Wohlwend A, Vassalli JD. Extracellular proteolysis in the adult murine brain. J Clin Invest 1993;92:679-685.
- Strickland S, Reich E, Sherman MI. Plasminogen activator in early embryogenesis: enzyme production by trophoblast and parietal endoderm. Cell 1976;9:231-240.
- Smokovitis A, Kok P, Astrup T. Tissue repair in rats in presence of locally applied tissue plasminogen activator. Exp Mol Pathol 1975;22:109-117.
- Aguilar S, Corominas JM, Malats N, Pereira JA, Dufresne M, Real FX, Navarro P. Tissue plasminogen activator in murine exocrine pancreas cancer: selective expression in ductal tumors and contribution to cancer progression. Am J Pathol 2004;165:1129-1139.
- Lee HY, Hwang IY, Im H, Koh JY, Kim YH. Non-proteolytic neurotrophic effects of tissue plasminogen activator on cultured mouse cerebrocortical neurons. J Neurochem 2007;101:1236-1247.
- Pittman RN, Ivins JK, Buettner HM. Neuronal plasminogen activators: cell surface binding sites and involvement in neurite outgrowth. J Neurosci 1989;9:4269-4286.
- Nicole O, Docagne F, Ali C, Margaill I, Carmeliet P, MacKenzie ET, Vivien D, Buisson A. The proteolytic activity of tissue-plasminogen activator enhances NMDA receptor-mediated signaling. Nat Med 2001;7:59-64.
- Peters S, Koh J, Choi DW. Zinc selectively blocks the action of N-methyl-D-aspartate on cortical neurons. Science 1987;236:589-593.
- Koh JY, Suh SW, Gwag BJ, He YY, Hsu CY, Choi DW. The role of zinc in selective neuronal death after transient global cerebral ischemia. Science 1996;272:1013-1016.
- Kim YH, Park JH, Hong SH, Koh JY. Nonproteolytic neuroprotection by human recombinant tissue plasminogen activator. Science 1999;284:647-650.
- Qian Z, Gilbert ME, Colicos MA, Kandel ER, Kuhl D. Tissue-plasminogen activator is induced as an immediate-early gene during seizure, kindling and long-term potentiation. Nature 1993;361:453-457.
- Baranes D, Lederfein D, Huang YY, Chen M, Bailey CH, Kandel ER. Tissue plasminogen activator contributes to the late phase of LTP and to synaptic growth in the hippocampal mossy fiber pathway. Neuron 1998;21:813-825.
- Pang PT, Teng HK, Zaitsev E, Woo NT, Sakata K, Zhen S, Teng KK, Yung WH, Hempstead BL, Lu B. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science 2004;306:487-491.
- Chen ZL, Strickland S. Neuronal death in the hippocampus is promoted by plasmin-catalyzed degradation of laminin. Cell 1997;91:917-925.
- Chen ZL, Indyk JA, Strickland S. The hippocampal laminin matrix is dynamic and critical for neuronal survival. Mol Biol Cell 2003;14:2665-2676.
- Wang X, Lee SR, Arai K, Lee SR, Tsuji K, Rebeck GW, Lo EH. Lipoprotein receptor-mediated induction of matrix metalloproteinase by tissue plasminogen activator. Nat Med 2003;9:1313-1317.
- Lee JY, Kweon HS, Cho E, Lee JY, Byun HR, Kim DH, Kim YH, Han PL, Koh JY. Upregulation of tPA/plasminogen proteolytic system in the periphery of amyloid deposits in the Tg2576 mouse model of Alzheimer's disease. Neurosci Lett 2007;423:82-87.
- Ledesma MD, Da Silva JS, Crassaerts K, Delacourte A, De Strooper B, Dotti CG. Brain plasmin enhances APP alpha-cleavage and Abeta degradation and is reduced in Alzheimer's disease brains. EMBO Rep 2000;1:530-535.
- Tucker HM, Kihiko M, Caldwell JN, Wright S, Kawarabayashi T, Price D, Walker D, Scheff S, McGillis JP, Rydel RE, Estus S. The plasmin system is induced by and degrades amyloid-beta aggregates. J Neurosci 2000;20:3937-3946.
- Hwang IY, Sun ES, An JH, Im H, Lee SH, Lee JY, Han PL, Koh JY, Kim YH. Zinc-triggered induction of tissue plasminogen activator by brain-derived neurotrophic factor and metalloproteinases. J Neurochem 2011;118:855-863.
- Tigges U, Welser-Alves JV, Boroujerdi A, Milner R. A novel and simple method for culturing pericytes from mouse brain. Microvasc Res 2012;84:74-80.
- Cort J, Liu Z, Lee G, Harris SM, Prickett KS, Gaeta LS, Andersen NH. Beta-structure in human amylin and two designer beta-peptides: CD and NMR spectroscopic comparisons suggest soluble beta-oligomers and the absence of significant populations of beta-strand dimers. Biochem Biophys Res Commun 1994;204:1088-1095.
- Kingston IB, Castro MJ, Anderson S. In vitro stimulation of tissue-type plasminogen activator by Alzheimer amyloid beta-peptide analogues. Nat Med 1995;1:138-142.
- Emmetsberger J, Mirrione MM, Zhou C, Fernandez-Monreal M, Siddiq MM, Ji K, Tsirka SE. Tissue plasminogen activator alters intracellular sequestration of zinc through interaction with the transporter ZIP4. J Neurosci 2010;30:6538-6547.
- Melchor JP, Pawlak R, Strickland S. The tissue plasminogen activator-plasminogen proteolytic cascade accelerates amyloid-beta (Abeta) degradation and inhibits Abeta-induced neurodegeneration. J Neurosci 2003;23:8867-8871.