Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2014; 23(2): 148-154
Published online June 30, 2014
https://doi.org/10.5607/en.2014.23.2.148
© The Korean Society for Brain and Neural Sciences
Suppression of miR-155 Expression in IFN-γ-Treated Astrocytes and Microglia by DJ-1: A Possible Mechanism for Maintaining SOCS1 Expression
Jong-hyeon Kim1,2,3,4, Ilo Jou1,2,4 and Eun-Hye Joe1,2,3,4*
1Department of Biomedical Sciences, Neuroscience Graduate Program, 2Department of Pharmacology, 3Department of Brain Science, 4Chronic Inflammatory Disease Research Center, Ajou University School of Medicine, Suwon 443-380, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-31-219-5062, FAX: 82-31-219-5069
e-mail: ehjoe@ajou.ac.kr
Previously, we reported that DJ-1, encoded by a Parkinson's disease (PD)-associated gene, inhibits expression of proinflammatory mediators in interferon-gamma (IFN-γ)-treated astrocytes and microglia through inhibition of STAT1 activation. Here, using microglia and astrocytes cultured from wild-type (WT) and DJ-1-knockout (KO) mouse brains, we examined how DJ-1 regulates suppressor of cytokine signaling 1 (SOCS1), a negative feedback regulator of STAT1 (signal transducer and activator of transcription) that is also induced by STAT1. We found that IFN-γ significantly increased SOCS1 mRNA expression in WT microglia and astrocytes, but not in KO cells, although STAT1 was highly activated in these latter cells. We further found that SOCS mRNA stability was decreased in DJ-1-KO cells, an effect that appeared to be mediated by the microRNA, miR-155. IFN-γ increased the levels of miR-155 in DJ-1-KO cells but not in WT cells. In addition, an miR-155 inhibitor rescued SOCS1 expression and decreased STAT1 activation in DJ-1-KO cells. Taken together, these results suggest that DJ-1 efficiently regulates inflammation by maintaining SOCS1 expression through regulation of miR-155 levels, even under conditions in which STAT1 activation is decreased.
Keywords: parkinson, DJ-1, SOCS1, miR-155, inflammation
The symptoms of Parkinson's disease (PD), the second leading neurodegenerative disease, are related to dopaminergic neuronal death in the substantia nigra pars compacta. Although most cases of PD are sporadic, a significant research effort has been devoted to studying the function of PD-associated genes to obtain insight into the onset and progression of PD. DJ-1, also known as PARK7 (Parkinson protein 7), is an early onset autosomal-recessive PD gene [1]. Although DJ-1 has been identified as an oncogene [2], it has diverse functions. For example, it stabilizes the transcription factor, Nrf2, which regulates expression of anti-oxidant enzymes [3], and modulates cell death through regulation of apoptosis-associated proteins [4, 5, 6]. DJ-1 also acts as a chaperone to suppress fibrillation of a-synuclein [7]. Recently, we and others reported that DJ-1 possesses anti-inflammatory functions [8, 9]. Our studies provided insight into the mechanism underlying this anti-inflammation role, showing that DJ-1 facilitates the interaction between STAT1 (signal transducer and activator of transcription) and its phosphatase SHP-1 (Src homology region 2 domain-containing phosphatase-1), and thereby inhibits STAT1 activation [9].
Inflammation, including brain inflammation, is a defense mechanism that provides protection against infection. However, excessive and long-lasting inflammation can be toxic to surrounding tissues, particularly in the brain where neurons regenerate poorly once damaged. Thus, not surprisingly, the extent and duration of brain inflammation is tightly regulated through diverse mechanisms [10, 11]. Suppressor of cytokine signaling (SOCS) family proteins, including SOCS1-7 and CIS (cytokine-inducible SH2-containing protein) [12, 13], are well-characterized negative feedback regulators of inflammation. Inflammatory stimulators, including interferon gamma (IFN-γ), lipopolysaccharide (LPS) and thrombin, among others, induce expression of SOCS family proteins as well as proinflammatory mediators [14, 15, 16, 17]. Furthermore, it appears that SOCS proteins and pro-inflammatory mediators are induced through the same signaling pathways and with a similar time course [13, 16, 17]. IFN-γ induces expression of SOCS1 and SOCS3 mRNA within a few hours through activation of the JAK/STAT pathway [12, 13]. In macrophages, CpG DNA induces SOCS1 and SOCS3 expression via mitogen-activated protein kinase (MAPK), particularly ERK (extracellular signal-regulated kinase) and p38 MAPK [18]. In brain microglia and astrocytes, thrombin triggers SOCS3 and CIS expression through protein kinase C-delta (PKCd) and reactive oxygen species (ROS), respectively [16, 17]. Notably, these signaling molecules play an important in mediating the expression of pro-inflammatory mediators in microglia and macrophages [19, 20, 21, 22, 23, 24, 25, 26]. In fact, SOCS1 and SOCS3, induced by activation of the JAK/STAT pathway, in turn inhibit JAK/STAT signaling, providing a feedback mechanism for curtailing the inflammation process [12, 13]. Accordingly, SOCS1-deficient cells and mice display hyper-responsiveness to inflammatory stimuli [27, 28,29], and mutation of SOCS3 exacerbates colitis [30].
Previously, we reported that DJ-1 exerts anti-inflammatory effects through inhibition of STAT1 activation [9]. These finding led us to ask how DJ-1 regulates SOCS1 expression, since SOCS1 expression is induced by STAT1 activation. Interestingly, we found that DJ-1 regulates SOCS1 expression through the SOCS1-specific microRNA, miR-155, a mechanism distinct from that by which it regulates proinflammatory mediators.
MATERIALS AND METHODS
DJ-1-KO mice, a generous gift from Dr. U. J. Kang (Chicago University, Chicago, IL, USA), were generated previously by deleting a 9.3-kb region of genomic DNA containing the first five exons and part of the promoter region of the DJ-1 gene [31].
Primary astrocytes and microglia were cultured from the forebrain of 1-day-old mice [9]. In brief, forebrains were removed into Dulbecco's modified Eagle medium (DMEM; Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT, USA) and triturated using a pipette. Dissociated cells were then plated in 75-cm2 T-flasks (0.5 hemisphere/flask) and cultured for 2~3 weeks. Microglia were detached from flasks by gently shaking, filtered through a nylon mesh to remove cell clumps, and incubated in DMEM containing 10% FBS. Astrocytes were harvested with 0.1% trypsin and incubated in DMEM containing 10% FBS. For activation of glial cells, cells were treated with 10 ng/ml recombinant murine IFN-γ (PeproTech, Rocky Hill, NJ, USA).
Total RNA was isolated using RNAzol B (iNtRON, Sungnam, Korea), and cDNA was prepared using Avian Myeloblastosis Virus reverse transcriptase (Promega, Madison, WI, USA) according to the manufacturers' instructions. TNF-α, SOCS1, and β-actin transcripts were measured by quantitative real-time polymerase chain reaction (qPCR) on a RotoGene thermocycler (Corbett Research, Sydney, Australia) using a 2× KAPA SYBR Fast Master Mix (Kapa Biosystems, Cape Town, South Africa) and the following primer pairs: TNF-α, 5'-GTAGCCCACGTCGTAGCAAA-3' (sense) and 5'-CCCTTCTCCAGCTGGGAGAC-3' (antisense); SOCS1, 5'-ACACTCACTTCCGCACCTTC-3' (sense) and 5'-CACGGAGTACCGGGTTAAGA-3' (antisense); β-actin, 5'-GCTCTGGCTCCTAGCACCAT-3' (sense) and 5'-GCCACCGATCCACACAGAGT-3' (antisense). microRNA was isolated using the miRNeasy Mini Kit (QIAGEN, Valencia, CA, USA), and cDNA was prepared using the miScript II RT Kit (QIAGEN), according to the manufacturer's instructions.
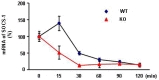
mRNA stability was measured by treating cells first with IFN-γ for 3 hours and then with 2 ng/ml actinomycin D (ActD; SIGMA-ALDRICH, St. Louis, MO, USA) for up to 120 minutes. The remaining SOCS1 transcript levels were measured by qPCR.
Levels of miR-155 and the housekeeping microRNA RNU were measured by qPCR using a miScript SYBR Green PCR Kit (QIAGEN) and a RotoGene thermocycler (Corbett Research). miR-155 and RNU primers were purchased from QIAGEN. The threshold cycle number was calculated for each gene and normalized to that of β-actin or RNU. The Δ-δ threshold cycle values for each gene are presented as relative fold induction.
Cells were washed with ice-cold phosphate-buffered saline (PBS) and lysed on ice in RIPA buffer (50 mM Tris-HCl pH 7.4, 1% NP-40, 0.25% Na-deoxycholate, 150 mM NaCl, 1 mM Na3VO4, and 1 mM NaF) containing protease inhibitor and phosphatase inhibitor cocktails (GenDEPOT, Barker, TX, USA). Lysates were centrifuged, and proteins in the supernatant were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF (polyvinylidene difluoride) membranes. Membranes were incubated with antibodies specific for phospho-Tyr-STAT1 (pY-STAT1; 1:1000; Cell Signaling Technology, Beverly, MA, USA) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:1000; Santa Cruz Biotechnology, CA, USA). After washing with PBS containing 0.05% Tween-20, membranes were incubated with peroxidase-conjugated secondary antibodies (1:10,000; Zymed, San Francisco, CA, USA), and immunoreactive proteins were visualized using an enhanced chemiluminescence system (Daeil Lab, Seoul, Korea).
DJ-1-KO astrocytes were maintained at 60%~80% confluence. Cells were transiently transfected with miR-155 inhibitor (QIAGEN; 5'-UUAAUGCUAAUUGUGAUAGGGGU-3') using a mixture of opti-MEM (Invitrogen, Carlsbad, CA, USA) and RNAiMAX transfection reagents (Invitrogen) for 6 hours, according to the manufacturer's instructions.
All data presented in this study are representative of at least three independent experiments. Data were analyzed using Student's t-tests.
RESULTS AND DISCUSSION
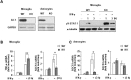
In our previous study, we reported that a DJ-1 deficiency increased expression of proinflammatory mediators in brain microglia and astrocytes through enhanced STAT1 activation [9]. Here, we examined whether DJ-1 enhanced the expression of the negative regulator of inflammation, SOCS1, since SOCS1 is also induced by STAT1 activation [12, 13]. First, we confirmed induction of DJ-1 and TNF-α expression and STAT1 activation by IFN-γ in microglia and/or astrocytes prepared from DJ-1-KO mice and WT mice (Fig. 1A, B). Interestingly, the expression patterns of SOCS1 were different from those of TNF-α. In keeping with the enhanced STAT1 activation observed in KO cells (Fig. 1A), IFN-γ induced TNF-α expression more strongly in KO microglia (Fig. 1B), as shown in our previous study [9], but induced SOCS1 expression more strongly in WT microglia and astrocytes than in KO cells (Fig. 1B, C). This latter effect appeared to be specific for the SOCS1 isoform since SOCS3 expression was not different between WT and KO cells (Fig. 1C). These results show that DJ-1 differentially regulates the expression of proinflammatory mediator(s) and SOCS1, inhibiting TNF-α expression and enhancing SOCS1 expression.
Next, we examined the mechanisms underlying the reduced SOCS1 mRNA expression in DJ-1-KO cells. An increase in mRNA levels could reflect increased transcriptional activation or enhanced mRNA stability. We excluded transcriptional activation since activation of STAT1, which positively regulates SOCS1 transcription [12, 13], was more strongly activated in DJ-1-KO cells (Fig. 1A). To determine whether SOCS1 mRNA levels were maintained through enhanced mRNA stability, we treated mixed cultures of astrocytes and microglia with IFN-γ to induce SOCS1, added actinomycin D (ActD), which inhibits mRNA transcription, and then measured SOCS1 mRNA levels. Although SOCS1 mRNA levels decreased in the presence of ActD in both WT and KO cells, the SOCS1 mRNA degradation rates were much faster in DJ-1-KO cells than in DJ-1-WT cells (Fig. 2). These results suggest that DJ-1 protects SOCS1 mRNA from degradation, providing a mechanism for maintenance of IFN-γ-induced SOCS1 mRNA levels in WT cells.
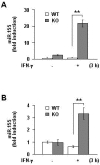
Several previous studies have reported that miR-155 down-regulates SOCS1 [32, 33, 34]. On the basis of these observations, we measured miR-155 levels in WT and KO microglia and astrocytes. Interestingly, IFN-γ increased miR-155 expression within 3 hours in KO microglia and astrocytes, but not in WT cells (Fig. 3).
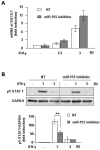
We further confirmed miR-155-mediated regulation of SOCS1 mRNA levels using an miR-155 inhibitor (see Materials and methods). DJ-1-KO astrocytes were transfected with an miR-155 inhibitor for 6 hours, and then treated with IFN γ. We found that the miR-155 inhibitor further increased IFN-γ-induced SOCS1 expression (Fig. 4A). We additionally examined whether miR-155 inhibition decreased STAT1 activation, consistent with the role of SOCS1 as a negative regulator of STAT1 activation. As expected, STAT1 activation by IFN-γ was less robust in cells transfected with the miR-155 inhibitor (Fig. 4B). Taken together, these results suggest that DJ-1 exerts an anti-inflammatory effect by maintaining SOCS1 expression through regulation of miR-155 expression.
Brain inflammation has been suggested as a risk factor for PD. Although the brain inflammation that accompanies acute brain injury does not appear to be toxic to neurons [11, 35], PD genes could alter this normal pattern. In this context, we and others have reported that both loss-of-function mutants of PD genes (PINK and DJ-1) and gain-of-function mutants of PD genes (a-synuclein and LRRK2) enhance inflammatory responses [9, 36, 37, 38]. Therefore, mutations of PD genes could cause excessive inflammatory responses in the injured brain.
SOCS1 is a well-known negative feedback inhibitor of STAT1 activation [12, 13]. Therefore, a decrease in SOCS1 expression would result in inefficient termination of STAT1-induced inflammation. IFN-γ induces SOCS1 expression in the same way that it induces proinflammatory mediators-via STAT1 activation [12, 13]. Moreover, DJ-1 facilitates the interaction between STAT1 and its phosphatase, SHP-1 [9], thereby inhibiting STAT1 activation and the expression of proinflammatory mediators. Thus, if there are no other mechanisms that regulated SOCS1 expression, DJ-1 would down-regulate SOCS1 expression since it inhibits STAT1 activation (Fig. 1A). However, in this study, we found that, although DJ-1 suppresses the expression of proinflammatory mediators through inhibition of STAT1 activation, it maintains SOCS1 expression through regulation of miR-155 levels, even under conditions in which STAT1 activation is decreased (Fig. 3). Collectively, these observations demonstrate a novel mechanism for controlling inflammation by DJ-1.
Most studies on neurodegenerative diseases, including PD, have focused on neurons because neuronal death is related to the appearance of symptoms. However, neuronal death could be caused by abnormally functioning astrocytes and microglia. Since PD-associated genes, including DJ-1, PINK1, parkin and LRRK2, are expressed in astrocytes and microglia [9, 39, 40, 41, 42], mutations in these genes could alter the function of these cells. To our knowledge, this study is the first to report that DJ-1 regulates expression of SOCS1 and miR-155. Accordingly, mutations in DJ-1 would alter the inflammatory responses of microglia and astrocytes, highlighting the importance of investigating astrocytic and microglial processes in studies of PD and other neurodegenerative diseases.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Bonifati V, Rizzu P, van Baren MJ, Schaap O, Breedveld GJ, Krieger E, Dekker MC, Squitieri F, Ibanez P, Joosse M, van Dongen JW, Vanacore N, van Swieten JC, Brice A, Meco G, van Duijn CM, Oostra BA, Heutink P. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003;299:256-259.

- Nagakubo D, Taira T, Kitaura H, Ikeda M, Tamai K, Iguchi-Ariga SM, Ariga H. DJ-1, a novel oncogene which transforms mouse NIH3T3 cells in cooperation with ras. Biochem Biophys Res Commun 1997;231:509-513.
- Clements CM, McNally RS, Conti BJ, Mak TW, Ting JP. DJ-1, a cancer- and Parkinson's disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc Natl Acad Sci U S A 2006;103:15091-15096.
- Fan J, Ren H, Jia N, Fei E, Zhou T, Jiang P, Wu M, Wang G. DJ-1 decreases Bax expression through repressing p53 transcriptional activity. J Biol Chem 2008;283:4022-4030.
- Junn E, Taniguchi H, Jeong BS, Zhao X, Ichijo H, Mouradian MM. Interaction of DJ-1 with Daxx inhibits apoptosis signal-regulating kinase 1 activity and cell death. Proc Natl Acad Sci U S A 2005;102:9691-9696.
- Ren H, Fu K, Wang D, Mu C, Wang G. Oxidized DJ-1 interacts with the mitochondrial protein BCL-XL. J Biol Chem 2011;286:35308-35317.
- Shendelman S, Jonason A, Martinat C, Leete T, Abeliovich A. DJ-1 is a redox-dependent molecular chaperone that inhibits alpha-synuclein aggregate formation. PLoS Biol 2004;2:e362.
- Waak J, Weber SS, Waldenmaier A, Görner K, Alunni-Fabbroni M, Schell H, Vogt-Weisenhorn D, Pham TT, Reumers V, Baekelandt V, Wurst W, Kahle PJ. Regulation of astrocyte inflammatory responses by the Parkinson's disease-associated gene DJ-1. FASEB J 2009;23:2478-2489.
- Kim JH, Choi DJ, Jeong HK, Kim J, Kim DW, Choi SY, Park SM, Suh YH, Jou I, Joe EH. DJ-1 facilitates the interaction between STAT1 and its phosphatase, SHP-1, in brain microglia and astrocytes: A novel anti-inflammatory function of DJ-1. Neurobiol Dis 2013;60:1-10.
- Yang MS, Min KJ, Joe E. Multiple mechanisms that prevent excessive brain inflammation. J Neurosci Res 2007;85:2298-2305.
- Jeong HK, Ji K, Min K, Joe EH. Brain inflammation and microglia: facts and misconceptions. Exp Neurobiol 2013;22:59-67.
- Krebs DL, Hilton DJ. SOCS proteins: negative regulators of cytokine signaling. Stem Cells 2001;19:378-387.
- Yasukawa H, Sasaki A, Yoshimura A. Negative regulation of cytokine signaling pathways. Annu Rev Immunol 2000;18:143-164.
- Song MM, Shuai K. The suppressor of cytokine signaling (SOCS) 1 and SOCS3 but not SOCS2 proteins inhibit interferon-mediated antiviral and antiproliferative activities. J Biol Chem 1998;273:35056-35062.
- Crespo A, Filla MB, Russell SW, Murphy WJ. Indirect induction of suppressor of cytokine signalling-1 in macrophages stimulated with bacterial lipopolysaccharide: partial role of autocrine/paracrine interferon-alpha/beta. Biochem J 2000;349:99-104.
- Ji KA, Yang MS, Jou I, Shong MH, Joe EH. Thrombin induces expression of cytokine-induced SH2 protein (CIS) in rat brain astrocytes: involvement of phospholipase A2, cyclooxygenase, and lipoxygenase. Glia 2004;48:102-111.
- Yang MS, Lee J, Ji KA, Min KJ, Lee MA, Jou I, Joe E. Thrombin induces suppressor of cytokine signaling 3 expression in brain microglia via protein kinase Cdelta activation. Biochem Biophys Res Commun 2004;317:811-816.
- Dalpke AH, Opper S, Zimmermann S, Heeg K. Suppressors of cytokine signaling (SOCS)-1 and SOCS-3 are induced by CpG-DNA and modulate cytokine responses in APCs. J Immunol 2001;166:7082-7089.
- Pyo H, Jou I, Jung S, Hong S, Joe EH. Mitogen-activated protein kinases activated by lipopolysaccharide and beta-amyloid in cultured rat microglia. Neuroreport 1998;9:871-874.
- Pyo H, Joe E, Jung S, Lee SH, Jou I. Gangliosides activate cultured rat brain microglia. J Biol Chem 1999;274:34584-34589.
- Ryu J, Pyo H, Jou I, Joe E. Thrombin induces NO release from cultured rat microglia via protein kinase C, mitogen-activated protein kinase, and NF-kappa B. J Biol Chem 2000;275:29955-29959.
- Kang J, Park EJ, Jou I, Kim JH, Joe EH. Reactive oxygen species mediate A beta(25-35)-induced activation of BV-2 microglia. Neuroreport 2001;12:1449-1452.
- Kang J, Yang M, Jou I, Joe E. Identification of protein kinase C isoforms involved in interferon-gamma-induced expression of inducible nitric oxide synthase in murine BV2 microglia. Neurosci Lett 2001;299:205-208.
- Kim OS, Park EJ, Joe EH, Jou I. JAK-STAT signaling mediates gangliosides-induced inflammatory responses in brain microglial cells. J Biol Chem 2002;277:40594-40601.
- Min KJ, Jou I, Joe E. Plasminogen-induced IL-1beta and TNF-alpha production in microglia is regulated by reactive oxygen species. Biochem Biophys Res Commun 2003;312:969-974.
- Min KJ, Pyo HK, Yang MS, Ji KA, Jou I, Joe EH. Gangliosides activate microglia via protein kinase C and NADPH oxidase. Glia 2004;48:197-206.
- He Y, Zhang W, Zhang R, Zhang H, Min W. SOCS1 inhibits tumor necrosis factor-induced activation of ASK1-JNK inflammatory signaling by mediating ASK1 degradation. J Biol Chem 2006;281:5559-5566.
- Hanada T, Yoshida H, Kato S, Tanaka K, Masutani K, Tsukada J, Nomura Y, Mimata H, Kubo M, Yoshimura A. Suppressor of cytokine signaling-1 is essential for suppressing dendritic cell activation and systemic autoimmunity. Immunity 2003;19:437-450.
- Chinen T, Kobayashi T, Ogata H, Takaesu G, Takaki H, Hashimoto M, Yagita H, Nawata H, Yoshimura A. Suppressor of cytokine signaling-1 regulates inflammatory bowel disease in which both IFNgamma and IL-4 are involved. Gastroenterology 2006;130:373-388.
- Suzuki A, Hanada T, Mitsuyama K, Yoshida T, Kamizono S, Hoshino T, Kubo M, Yamashita A, Okabe M, Takeda K, Akira S, Matsumoto S, Toyonaga A, Sata M, Yoshimura A. CIS3/SOCS3/SSI3 plays a negative regulatory role in STAT3 activation and intestinal inflammation. J Exp Med 2001;193:471-481.
- Chen L, Cagniard B, Mathews T, Jones S, Koh HC, Ding Y, Carvey PM, Ling Z, Kang UJ, Zhuang X. Age-dependent motor deficits and dopaminergic dysfunction in DJ-1 null mice. J Biol Chem 2005;280:21418-21426.
- Jiang S, Zhang HW, Lu MH, He XH, Li Y, Gu H, Liu MF, Wang ED. MicroRNA-155 functions as an OncomiR in breast cancer by targeting the suppressor of cytokine signaling 1 gene. Cancer Res 2010;70:3119-3127.
- Wang P, Hou J, Lin L, Wang C, Liu X, Li D, Ma F, Wang Z, Cao X. Inducible microRNA-155 feedback promotes type I IFN signaling in antiviral innate immunity by targeting suppressor of cytokine signaling 1. J Immunol 2010;185:6226-6233.
- Cardoso AL, Guedes JR, Pereira de Almeida L, Pedroso de Lima MC. miR-155 modulates microglia-mediated immune response by down-regulating SOCS-1 and promoting cytokine and nitric oxide production. Immunology 2012;135:73-88.
- Jeong HK, Ji KM, Kim B, Kim J, Jou I, Joe EH. Inflammatory responses are notsufficient to cause delayed neuronal death in ATP-induced acute brain injury. PLoS One 2010;5:e13756.
- Kim J, Byun JW, Choi I, Kim B, Jeong HK, Jou I, Joe E. PINK1 Deficiency Enhances Inflammatory Cytokine Release from Acutely Prepared Brain Slices. Exp Neurobiol 2013;22:38-44.
- Croisier E, Moran LB, Dexter DT, Pearce RK, Graeber MB. Microglial inflammation in the parkinsonian substantia nigra: relationship to alpha-synuclein deposition. J Neuroinflammation 2005;2:14.
- Su X, Maguire-Zeiss KA, Giuliano R, Prifti L, Venkatesh K, Federoff HJ. Synuclein activates microglia in a model of Parkinson's disease. Neurobiol Aging 2008;29:1690-1701.
- Kim B, Yang MS, Choi D, Kim JH, Kim HS, Seol W, Choi S, Jou I, Kim EY, Joe EH. Impaired inflammatory responses in murine Lrrk2-knockdown brain microglia. PLoS One 2012;7:e34693.
- Choi I, Kim J, Jeong HK, Kim B, Jou I, Park SM, Chen L, Kang UJ, Zhuang X, Joe EH. PINK1 deficiency attenuates astrocyte proliferation through mitochondrial dysfunction, reduced AKT and increased p38 MAPK activation, and downregulation of EGFR. Glia 2013;61:800-812.
- Ledesma MD, Galvan C, Hellias B, Dotti C, Jensen PH. Astrocytic but not neuronal increased expression and redistribution of parkin during unfolded protein stress. J Neurochem 2002;83:1431-1440.
- Solano RM, Casarejos MJ, Menéndez-Cuervo J, Rodriguez-Navarro JA, García de Yébenes J, Mena MA. Glial dysfunction in parkin null mice: effects of aging. J Neurosci 2008;28:598-611.