Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Review Article
Exp Neurobiol 2016; 25(5): 205-221
Published online October 31, 2016
https://doi.org/10.5607/en.2016.25.5.205
© The Korean Society for Brain and Neural Sciences
Optogenetic and Chemogenetic Approaches for Studying Astrocytes and Gliotransmitters
Juwon Bang, Hak Yeong Kim and Hyosang Lee*
Department of Brain and Cognitive Sciences, DGIST, Daegu 42988, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-53-785-6147, FAX: 82-53-785-6109
e-mail: hyosang22@dgist.ac.kr
Abstract
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
The brain consists of heterogeneous populations of neuronal and non-neuronal cells. The revelation of their connections and interactions is fundamental to understanding normal brain functions as well as abnormal changes in pathological conditions. Optogenetics and chemogenetics have been developed to allow functional manipulations both
Keywords: optogenetics, chemogenetics, astrocytes, channelrhodopsin, DREADD, gliotransmitter
INTRODUCTION
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Astrocytes are the most abundant population of non-neuronal cells in the brain. It is well known that they provide structural and metabolic support for neuronal networks, but a growing body of evidence indicates that they also play an active role in modulating neuronal activity. Astrocytes make close contact with perisynaptic regions, forming a functional structure called the "tripartite synapse," together with presynaptic and postsynaptic nerve terminals [1,2,3]. Indeed, one astrocyte in the hippocampus makes contact with tens of thousands of synapses [4].
It is well established that astrocytes clear away excessive neurotransmitters and ions released from synaptic clefts through uptake via specific transporters and channels. For example, astrocytes remove excess extracellular glutamate using sodium-dependent glutamate transporters, such as the glutamate aspartate transporter and glutamate type 1 transporter [5,6,7,8,9]. Excessive glutamate is cytotoxic to neurons, causing an influx of calcium that far exceeds physiological levels and triggering the activation of enzymes and signaling proteins that are detrimental to neurons [10,11,12].
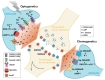
Evidence from a number of studies indicates that astrocytes release signaling molecules, the so-called "gliotransmitters," such as glutamate, GABA, D-serine, and ATP, into the extracellular milieu in response to extracellular stimuli (Fig. 1) [13,14,15,16,17,18]. Gliotransmitter release often results from the activation of G protein-coupled receptors (GPCRs) that trigger downstream signaling cascades in astrocytes, including phospholipase C, adenylate cyclase, inositol 1,4,5-trisphosphate (IP3), and cause an intracellular calcium increase [19,20,21]. Gliotransmitters facilitate or inhibit the excitability and synaptic transmission of neighboring neurons, and the outcome of their release is dependent on the site of action and types of activated receptors [3,13,14,22,23,24]. Through the use of volume-sensitive organic anion channels, gap junction hemichannels, P2X7, Bestrophin-1, and reverse-orientation plasma membrane glutamate transporters, diverse mechanisms for gliotransmitter release have been identified in astrocytes, including calcium-dependent exocytotic vesicular release as well as non-exocytotic mechanisms [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. Although accumulating evidence suggests a coupling between various intracellular changes in astrocytes, such as intracellular calcium increase and gliotransmitter release, there is also evidence against this view; thus, the mechanisms underlying astrocyte-neuronal communication are still debated [40,41,42,43,44,45]. In this review, we present recent studies that have using optogenetic and chemogenetic approaches to explore the function of astrocytes and gliotransmitters.
OPTOGENETICS AND CHEMOGENETICS
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Optogenetics is a novel biological technique based on a variety of light-sensitive proteins called opsins, which include microbial ion channels and ion pumps as well as engineered GPCRs (Fig. 1) [46,47]. Following absorption of a specific wavelength of light, an opsin undergoes a conformational change that triggers diverse cellular changes in opsin-expressing cells. Some of the opsins induce the translocation of ions, and others activate intracellular signaling cascades, such as G protein-mediated signaling. Since most of these opsins do not exist in experimental-model organisms, and photostimulation itself has a negligible effect on cells and tissues, optogenetics has instead been used as a powerful experimental tool to manipulate specific populations of cells both
Opsins have been modified to generate mutants and chimeric proteins with diverse features, including their intracellular effects, optimal wavelengths of light for activation, and temporal dynamics in activation and inactivation; thus, they offer great flexibility in designing experiments and conducting more refined manipulations [47,48].
Channelrhodopsin-2 (ChR2), originally identified in green algae, is a cation channel that becomes permeable to positively charged ions such as proton and sodium when it is stimulated with blue light [49,50,51]. When it is expressed in neurons, photostimulation elicits an influx of cations, which causes depolarization and the firing of action potentials in the stimulated cells [46]. An influx of protons though ChR2 can also acidify the cytosolic compartment of photostimulated cells [51,52]. In the study of neurons, the frequency and duration of neuronal spiking can be easily controlled using variants of ChR2, such as ChR2(H134R), ChR2(C128S), ChETA, and step function opsin (SFO) [53,54,55,56,57]. For example, ChETA can drive ultrafast spiking up to 200 Hz, and SFO can elicit prolonged, bi-stable, sub-threshold depolarization of membranes [55,56]. Some light-gated cation channels, such as calcium-translocating channelrhodopsin (CatCh) and LiGluR, are more permeable to calcium than ChR2, and therefore they have been preferentially used in studies exploring the role of intracellular calcium [58,59,60,61]. LiGluR is a mutated ionotropic glutamate receptor 6 containing its ligand attached to an optically switchable tether called maleimide-azobenzene-glutamate [61,62].
Halorhodopsin is an opsin identified from Archaea which, when stimulated with yellow light, pumps chloride ions into cells [63,64,65,66,67,68,69]. When halorhodopsin is expressed in neurons, photostimulation promotes an influx of chloride ions that results in hyperpolarization and the inhibition of the firing of action potentials in the stimulated cells. Archaerhodopsins, such as Arch and ArchT, are light-driven outward proton pumps that inhibit the firing of action potentials during photostimulation when they are expressed in neurons; the efflux of protons can also cause alkalization of the cytosol [70,71,72]. Finally, optoXRs, such as optoα1AR and optoβ2AR, are chimeric GPCRs in which the intracellular loops of rhodopsin are replaced with those of other GPCRs, such as adrenergic receptors and dopamine receptors [73,74]. Photostimulation can initiate diverse intracellular signaling cascades in target cells, depending on the type of G protein replacing the intracellular loops of rhodopsin. Thus, these opsins enable the acute activation of different GPCR signaling pathways in cultured cells and animals.
Chemogenetics is based on engineered proteins, such as GPCRs and ligand-gated ion channels, that are no longer responsive or only very weakly responsive to their endogenous ligands but strongly respond to synthetic chemical ligands that are otherwise biologically inert [75,76,77]. For example, hM3Dq, one of the designer receptors exclusively activated by designer drugs (DREADDs), is generated by multiple cycles of randomized mutagenesis of the human M3 muscarinic receptor, which is linked to the Gq protein [78]. It is neither sensitive to the endogenous muscarinic acetylcholine receptor ligand acetylcholine nor is it constitutively active, but it is strongly activated in response to a synthetic ligand, clozapine-N-oxide (CNO), with nanomolar potency [75,76]. In response to CNO, hM3Dq can induce an enhancement of neuronal excitability that can lead to burst-like firing [79,80,81,82,83]. Thus, it is one of the most frequently used chemogenetic tools to activate neurons.
Another DREADD molecule, hM4Di, is a mutant of the Gi-coupled human M4 muscarinic receptor that responds to CNO [75,80,84]. Upon an application of the chemical agonist, hM4Di activates the Gβγ subunit of the Gi protein, which then stimulates G protein inwardly rectifying potassium channels (GIRK), causing an efflux of potassium and a resulting robust hyperpolarization when it is expressed in neurons [85,86]. Thus, hM4Di has been used to silence spontaneous and depolarization-evoked neuronal firing [75].
Optogenetics and chemogenetics require the expression of genetically encoded effectors in target cells. This goal is often attained by injecting a virus (e.g., adeno-associated virus (AAV) or lentivirus) that encodes an effector into a target region in the brain or other tissue. Alternatively, the effector can be expressed as a transgene in a genetically engineered mouse line. By using cell type-specific promoters, such as the astrocyte-specific glial fibrillary acid protein (GFAP) promoter, the effector's expression can be restricted to a specific population (or more than one population) of cells [87,88,89]. An intersectional strategy based on a combination of specific promoters and genetic tools, such as Cre- and flippase-mediated recombination, can further restrict the effector expression to specific subpopulations [90,91]. In addition, other genetic tricks, such as the use of tetracycline-dependent transcriptional regulation, have been used to achieve temporal control as well as amplification of effector expression [42,92,93].
Optogenetics and chemogenetics have different features that make them well suited for different applications. Optogenetics can deliver photostimulation directly to target cells and manipulate cellular activity acutely and reversibly. In contrast, chemogenetics is ideal for a prolonged manipulation of cellular activity in the range of minutes to days, depending on the route of ligand delivery and the pharmacokinetic properties of the synthetic ligand(s) used. Optogenetics is excellent in generating spiking patterns that mimic the endogenous firing responses of neurons by using a pulse generator that produces lights with different frequencies and pulse durations. In addition, photostimulation can be delivered to different subcellular locations such as the soma and nerve terminals, a useful feature for studying neuronal circuits in the brain [47]. On the other hand, chemogenetics is less invasive in experimental animals and hampers animal behaviors only marginally, if at all, because it requires neither the installation of a fiber-optic cable within the brain nor a connection of the cable to a light source, such as a laser or a light-emitting diode (LED). Furthermore, some synthetic ligands for chemogenetics, such as CNO, can be delivered via the animal's water and/or food as well as by systemic injection, permitting the delivery of the ligand with minimal disturbance of the animals, particularly in the case of chronic manipulation [94,95,96]. This minimal invasiveness can be a strong advantage in the clinical application of chemogenetics.
Optogenetic and chemogenetic techniques have been most frequently used to investigate neuronal circuits, but they also have been used to study non-neuronal cells in the brain and peripheral tissues. In the following section, we will summarize the approaches and findings of recent studies that have employed these techniques to reveal the function of astrocytes (Table 1 and 2).
OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Studies using primary astrocytes and immortal astrocyte cell lines have shown that optogenetic stimulation can elicit an elevation of intracellular calcium and subsequent release of gliotransmitters that can activate adjacent astrocytes as well as neurons. For example, Berlinguer-Palamini et al. have reported that photostimulation of ChR2-expressing primary astrocytes using a LED can elicit an intracellular calcium increase and electrophysiological responses not only in the stimulated cells but also in co-cultured astrocytes and neurons that do not express ChR2 [97]. The calcium response in ChR2-negative cells is suppressed by the application of antagonists of N-methyl-D-aspartate (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors in the bath solution, suggesting that the response in ChR2-negative cells is mediated by glutamate released from ChR2-expressing astrocytes.
Similar results have been obtained in another study, in which the coupling has been demonstrated between an intracellular calcium increase in astrocytes and glutamate release [98]. Li et al. have performed fluorescent calcium imaging in primary cultures of mouse cortical astrocytes and shown that photostimulation of astrocytes expressing LiGluR can elicit calcium transients not only in stimulated LiGluR-expressing cells but also in neighboring astrocytes that do not express the opsin. The calcium response in LiGluR-negative astrocytes is affected by antagonists of glutamate receptors, but not by a gap junction blocker or an antagonist of extracellular ATP signaling, suggesting the involvement of glutamate in the communication between astrocytes. Further experiments have shown that the calcium transients in LiGluR-negative astrocytes are inhibited by an anion channel blocker but are unaffected by an inhibitor of V-ATPase, which blocks exocytosis, suggesting that LiGluR-evoked glutamate release is mediated by anion channels [42,99,100,101,102,103,104,105].
In an attempt to investigate whether intracellular ionic alteration in astrocytes triggers gliotransmitter release, Ono and coworkers have co-cultured an astrocytic cell line and a neuronal cell line; in response to photostimulation, the ChR2-expressing astrocytes exhibited diverse cellular changes, including an increase in intracellular sodium and calcium, intracellular acidification, glutamate release, and inhibition of proliferation [106]. A short period of photostimulation (for minutes) elicited calcium transients in the co-cultured ChR2-negative neurons, whereas a long period of stimulation for several days resulted in apoptotic responses in the neurons. Both responses were blocked by an AMPA receptor antagonist. Thus, this study has demonstrated that activation of astrocytes releases glutamate which, in turn, provokes an intracellular calcium increase and cytotoxic cell death.
To mimic GPCR-mediated signaling events occurring in astrocytes in response to extracellular neurotransmitters and neuromodulators, Figueiredo et al. have expressed GPCR-based opsins, such as optoα1AR and optoβ2AR, in astrocytes to activate Gq- and Gs-mediated signaling cascades, respectively [107]. Photostimulation elicited an intracellular calcium increase in astrocytes expressing either opsin, which was blocked by apyrase, an enzyme hydrolyzing extracellular ATP, as well as by pharmacological blockers for the corresponding intracellular signaling cascade, such as inhibitors of phospholipase C and adenylate cyclase; these data indicate that a large portion of the calcium rise that was evoked by the activation of either opsin was a result of the autocrine action of extracellular ATP [108]. This study has demonstrated that GPCR-based opsins can be effectively used in the study of astrocytic GPCR-mediated signaling.
OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Tetracycline-dependent expression, using the so-called Tet-Off system, has been employed in several studies to generate transgenic mouse models expressing an opsin. In the Tet-Off system, the tetracycline-controlled transcriptional activator (tTA) binds to a tTA-responsive promoter sequence (tetO) to induce the expression of a downstream gene [109]. When bound to the tetracycline derivative doxycycline, tTA undergoes a conformational change that prevents tTA from binding to tetO, inhibiting the transcription of a target gene. Thus, this system enables a reversible control of gene expression produced by treatment with doxycycline. Tanaka and coworkers have generated a mouse line by knocking in a transgene cassette encoding tetO-driven ChR2(C128S) downstream of a housekeeping gene, β-actin, to obtain a high level expression [110]. The knockin mice have been crossed to tTA driver lines in which tTA is driven by cell type-specific promoters, the Mlc1, PLP, and Iba-1 promoters, to induce ChR2(C128S) expression in astrocytes, oligodendrocytes, and microglia, respectively.
The double-transgenic mice containing Mlc1-driven tTA and tetO-driven ChR2(C128S) have been used to reveal the role of Bergman glia (BG), a specialized subtype of astrocytes in the cerebellum, in modulating the activity of Purkinje neurons. First, photostimulation of acute brain slices prepared from the transgenic mice was found to elicit current responses from ChR2-expressing BG, suggesting that ChR2 is expressed in glial cells to a level sufficient for electrophysiological responses [70,110,111]. Second, photostimulation of the cerebellum using a fiber-optic cable installed above the skull, to avoid the generation of injury-induced reactive gliosis, was found to be sufficient to evoke an induction of a surrogate marker for cellular activation, c-fos, in ChR2-expressing BG [110]. Third, photostimulation of ChR2-expressing BG in acute cerebellar slices was shown to be sufficient to trigger glutamate release and firing of nearby Purkinje cells (PCs), resulting in long-term plasticity between parallel fibers and PCs [111]. Finally,
Beppu et al. have recently demonstrated using the same mouse line that neuronal damage in the mouse model of ischemia can be exacerbated by optogenetically induced acidosis and attenuated by alkalization of the cytosolic compartment of astrocytes [11,70]. Under ischemic conditions, such as deprivation of oxygen and glucose, cerebellar BG exhibited intracellular acidosis and glutamate release, followed by an inward excitatory current in the surrounding PCs. The response in PCs was inhibited by a cocktail of glutamate receptor and transporter blockers, suggesting the involvement of glutamate in the interaction between BG and PCs. Acidosis induced in BG by optogenetic stimulation of ChR2(C128S) was sufficient to evoke an inward excitatory current in the adjacent PCs. The response in PCs was inhibited by a non-competitive AMPA and kainate receptor antagonist, confirming the involvement of glutamate in the signaling between BG and PC. In contrast, an efflux of proton from BG produced by optogenetic stimulation of a light-gated outward proton pump, ArchT, led to a reduction in the inward currents in the PCs elicited by the deprivation of oxygen and glucose. Furthermore,
Using the same mouse line, Masamoto et al. have demonstrated that
OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
A number of studies have employed virally mediated expression of opsins to manipulate astrocytes, despite the possibility of inducing reactive gliosis as a result of viral infection [116]. This approach appears to be as successful as using transgenic mouse lines. For example, an AAV encoding GFAP promoter-driven ChR2 has been used to reveal a causal relationship between the activity of astrocytes and visual processing in the primary visual cortex (V1) [117]. Although a previous study had shown that astrocytes in the visual cortex respond to visual sensory stimuli, their roles had not been clearly determined because of the difficulty in selectively manipulating astrocytes among the heterogeneous populations of cells in the region. Perea et al. reported that
Optogenetic manipulations have revealed the involvement of other signaling molecules released by astrocytes, such as extracellular ATP and L-lactate, in modulating the activity of neurons in the brainstem. Gourine et al. have reported that astrocytes in the ventral surface of the medulla oblongata (VS) are exquisitely pH-sensitive [16]. In response to a decrease in pH in anesthetized rats, astrocytes residing near the VS exhibited an intracellular calcium increase. Furthermore, a decrease in pH in brainstem slices elicited a sustained ATP release in the VS region, as well as extracellular ATP-dependent calcium responses in VS astrocytes. To mimic pH-elicited calcium excitation in astrocytes, an AAV encoding enhanced GFAP promoter-driven ChR2(H134R) was injected into the brainstem. In organotypic brainstem slices, photostimulation elicited not only calcium transients in ChR2-expressing astrocytes but also long-lasting depolarization in adjacent chemo-sensitive neurons in the retrotrapezoid nucleus (RTN). RTN neurons have been found to play an important role in monitoring glucose concentrations, pH, and partial pressure of CO2 [118,119]. Either apyrase or an antagonist of extracellular ATP receptor blocked the response of the RTN neurons, suggesting that extracellular ATP mediates the interaction between astrocytes and adjacent neurons. Finally,
The same group of researchers has investigated the astrocytic modulation of norepinephrine (NE) release in the locus coeruleus (LC) using an AAV encoding GFAP promoter-driven ChR2(H134R) [120]. Evidence existed to suggest that L-lactate is involved in the process, but the exact mechanism was unclear [121,122,123]. Photostimulation of ChR2-expressing astrocytes in organotypic cultured brain slices elicited delayed depolarization and increased firing rates in norepinephrine (NE)-ergic neurons. Pharmacological interventions that reduce the level of L-lactate suppressed light-induced depolarization of NEergic neurons, suggesting that astrocytes activate NEergic neurons via L-lactate. Indeed, the application of L-lactate to brain slices provoked similar electrophysiological responses in NEergic neurons and caused NE release from the activated neurons. Finally, optogenetic activation of astrocytes using either optoβ2AR or ChR2(H134R) was sufficient to trigger NE release. Thus, this study clearly demonstrated that activated astrocytes in the LC release L-lactate, which then triggers NE release from NEergic neurons.
A similar viral approach was used by Gradinaru et al. to examine whether local astrocytes contribute to the therapeutic effect of deep-brain stimulation (DBS) delivered to the subthalamic nucleus (STN) to relieve tremor in Parkinson's disease [124]. To deliver photostimulation and measure neuronal activity from a parkinsonian rodent model, optrode recordings were performed in anesthetized rats, in which 6-hydroxydopamine (6-OHDA) had been unilaterally injected into the right medial forebrain bundle to cause a loss of nigral dopaminergic cells. 6-OHDA-treated animals displayed rotations ipsilateral to the lesion as a result of specific deficits in contralateral limb function, which became more obvious when amphetamine was administered to the subjects to increase locomotion. This study revealed that photostimulation of ChR2-expressing astrocytes in the STN can reversibly inhibit firing of STN neurons in 6-OHDA-treated animals; this treatment, however, failed to cause any changes in pathological motor behavior in parkinsonian rats, suggesting that astrocytes are unlikely to be critical players in the DBS-elicited effects.
Optogenetic manipulation has been used in two recent studies to examine the role of astrocytes in sleep. Pelluru et al. have reported that optogenetic activation of the posterior hypothalamus using ChR2(H134R) promotes both rapid and non-rapid eye movement sleep [125]. On the other hand, Yamashita et al. have reported that
In a very recent study reported by Poskanzer and Yuste, the role of neocortical astrocytes in the control of cortical circuit functions was examined using
CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Among diverse chemogenetic effectors, DREADDs, such as hM3Dq, have been used most frequently in studies focusing on astrocytes. As in optogenetic approaches, astrocyte-specific expression of chemogenetic proteins has been achieved by using viral and transgenic delivery in combination with astrocyte-specific promoters such as the GFAP and Mlc1 promoters.
To manipulate Gq-coupled receptor signaling in astrocytes, Fiacco et al. generated a bi-transgenic mouse line encoding GFAP promoter-driven tTA and tetO promoter-driven Mas-related G protein-coupled receptor A1 (MrgprA1) to express GPCR selectively in astrocytes [42,128]. MrgprA1 can be activated by RF amides, such as a peptide Phe-Leu-Arg-Phe amide (FLRFα) [129]. Since endogenous MrgprA1 is specifically expressed in dorsal root ganglion neurons but not in the brain, this protein is a useful molecular tool for manipulating neurons and glia in the brain when it is exogenously expressed in these cells. An infusion of FLRFα into acute hippocampal slices prepared from the transgenic mice elicited a robust calcium increase in widespread astrocytes, suggesting that MrgprA1 is able to activate the Gq-coupled intracellular signaling pathway. It is particularly interesting that such a widespread calcium rise in astrocytes failed to affect neuronal activity in the same slices. This finding is at odds with other studies, questioning the hypothesis that an astrocytic calcium increase causes the release of gliotransmitters which, in turn, activate nearby neurons [20,130,131,132,134,135,136]. In a follow-up study using MargprA1 mice together with mice lacking inositol 1,4,5-trisphosphate receptor 2 (IP3 R2), the astrocyte-specific IP3 receptor isoform, the same group of researchers further confirmed that activation of Gq protein-coupled signaling affects neither spontaneous excitatory postsynaptic currents nor the induction and maintenance of long-term potentiation in CA1 hippocampal neurons [40,137]. In another study performed by the same group of researchers, Bonder and McCarthy reported that hM3Dq can be selectively expressed in astrocytes by injecting AAV incorporating Cre-dependent hM3Dq into the visual cortex of transgenic mice encoding GFAP promoter-driven Cre, and they have used this system to investigate whether the astrocytic calcium elevation triggers vasodilation and a change in local blood flow in the cortex [41]. Activation of hM3Dq with CNO was sufficient to increase the intracellular calcium level in astrocytes but not to alter the basal blood flow in the visual cortex. The study also reported the absence of a temporal correlation between the astrocytic calcium increase and the change in cortical blood flow following either visual stimulation or a startle-evoking air puff. Furthermore, genetic deletion of IP3 R2 did not affect neurovascular coupling, suggesting that Gq signaling and IP3-dependent calcium elevation in astrocytes do not mediate vasodilation in the visual cortex.
The GFAP-driven MrgprA1 mouse line was used in a more recent study of Cao and coworkers to investigate the role of astrocytic ATP release in depression-like behaviors [138]. Application of the MrgprA1 agonist FLRFα elicited not only a robust increase in intracellular calcium in MrgprA1-expressing primary astrocytes but also a 2.5-fold increase in the ATP concentration in the culture medium. Furthermore, MrgprA1 mice infused with FLRFα into the cerebral ventricle exhibited a substantial reduction in depression-like behavior elicited in the murine paradigm of chronic social-defeat stress. Together with other results reported in the study, these findings suggest that endogenous ATP released from astrocytes can induce antidepressant-like behavior. Whether intracellular calcium increases in astrocytes can affect the activity of neighboring neurons was untested in the study.
Several other transgenic mouse lines expressing DREADDs have been reported. For instance, Sweger et al. developed a mouse line expressing an engineered k-opioid receptor (Ro1) in GFAP-expressing astrocytes by crossing transgenic mice encoding GFAP promoter-driven tTA mice with another line encoding tetO promoter-driven Ro1 on the background of a genetic deletion of the endogenous k-opioid receptor (KOR) [139]. Ro1 is a Gi-coupled GPCR that is insensitive to endogenous ligands of KOR, such as dynorphin, but highly sensitive to a synthetic ligand of the k-opioid receptor, spiradoline [140,141]. Ro1-expressing transgenic mice developed hydrocephalus and accumulation of cerebrospinal fluid in the ventricular system, even in the absence of a synthetic ligand, suggesting that Ro1 is constitutively active in this mouse model; unfortunately, this constitutive activity negates one of main features of chemogenetics, its temporal controllability, and limits the use of this model.
The same group of researchers has described another transgenic mouse line expressing GFAP promoter-driven hM3Dq [142]. Systemic treatment of these mice with CNO elicited a number of physiological changes that are controlled by the autonomic nervous system, including cardiovascular function, saliva formation, and homeostasis of body temperature. Furthermore, hM3Dq-expressing mice receiving CNO exhibited substantial changes in activity-related behaviors and motor coordination. Thus, these findings indicate the critical role of astrocytes in a broad range of basic physiological functions. Interestingly, the physiological and behavioral changes were unaffected by genetic deletion of IP3 R2, suggesting that astrocytic IP3-mediated calcium increase is dispensable for hM3Dq-elicited responses.
The GFAP promoter-driven hM3Dq mice have been used to study glial cells outside of the brain. McClain et al. examined the potential role of the enteric glia, which are astrocyte-like peripheral glial cells surrounding enteric neurons in the gut [143]. An application of CNO to the ileal and colonic myenteric plexus prepared from transgenic mice not only elicited and intracellular calcium increase in astrocytes but also triggered contraction of the ileum and colon to a degree similar to that elicited by direct stimulation of smooth muscle or electrical stimulation of enteric neurons. The contraction was abolished by the application of tetradotoxin, indicating the involvement of neuronal activation in the process. These findings have demonstrated that astrocytes in the gut play a critical role in the contractions of intestinal smooth muscle. The mechanism by which activation of Gq-coupled receptor in astrocytes leads to activation of enteric neurons remains unknown.
Recently, Sciolino et al. have reported two new transgenic mouse lines expressing hM3Dq, depending on Cre and flippase-mediated recombination [144]. When crossed to Cre or FLP driver lines, the new mouse lines permit the selective expression of hM3Dq in a population of cells that express either Cre or flippase. In addition, the intersectional strategy involving both Cre- and flippas-edependent recombination further restricts hM3Dq expression in a specific subpopulation. This group reported that a systemic application of CNO to mice expressing hM3Dq in GFAP-expressing cells elicits hypothermia, confirming the efficacy of the new mouse line by reproducing the previous finding [142].
Finally, the chemogenetic approach has also been applied to reveal the function of Gs-coupled signaling in longterm memory in normal animals and the Alzheimer animal model. For example, double-transgenic mice encoding GFAP promoter-driven tTA and tetO-driven Rs1 have been generated to acutely modulate Gs-coupled receptor activity [145]. Rs1 is the human Gs-coupled 5-HT4b serotonin receptor with a point mutation that renders this receptor insensitive to serotonin but highly sensitive to a synthetic ligand, GR-125487 [146]. Activation of Gs signaling by systemic delivery of GR-125487 impairs the performance of transgenic mice in the Morris water maze as well as a novel object-recognition task. Orr et al. found that Rs1 is constitutively active in this mouse model, driving the Gs signaling pathway even in the absence of the synthetic ligand.
CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Thus far, only a small number of studies focusing on glia have used a virally mediated method to achieve the expression of chemogenetic proteins. For example, an AAV encoding GFAP promoter-driven hM3Dq or hM4Di has been stereotactically injected into the arcuate nucleus of the mouse brain to investigate the potential role of medial basal hypothalamic astrocytes in regulating food intake [147]. In the feeding assay, hM3Dq-expressing mice receiving CNO exhibited a significant reduction in both baseline feeding and ghrelin-elicited hyperphagia, whereas hM4Di-expressing mice receiving CNO showed substantially enhanced and prolonged ghrelin-evoked feeding [148,149]. In contrast, following the CNO treatment, leptin-induced anorexia was facilitated in hM3Dq-expressing mice but suppressed in hM4Di-expressing mice [150,151,152,153]. Thus, this study employing two different chemogenetic actuators that recruit different downstream signaling molecules clearly demonstrated that astrocytes in the arcuate nucleus exert bi-directional regulation of food consumption.
An AAV virus expressing GFAP promoter-driven hM3Dq was injected into the rat nucleus accumbens core (NAcore) in two recent studies in order to investigate the contribution of glial cells and extracellular glutamate to substance abuse and motivation [154,155]. Bull et al. reported that an application of CNO elicited an elevation of the intracellular calcium level in hM3Dq-expressing primary astrocytes and a decrease in motivation for self-administration of ethanol after 3 weeks of abstinence [154]. Scofield et al. showed that intracranial or systemic administration of CNO triggered an increase in extracellular glutamate in the NAcore [155]. Furthermore, hM3Dq-expressing rats receiving intraperitoneal CNO exhibited a significant reduction in the cueinduced reinstatement of cocaine seeking [155].
CONCLUDING REMARKS
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
Neurons have always been a main focus of brain research, and non-neuronal cells that make up the majority of brain cells, such as glial cells, have not received much attention until recently. Studies of glia have revealed that they do not merely provide food and support to neurons; rather, they play an important role in brain function. In order to understand astrocytic function, it is critical to be able to control their intracellular activity in a native context. Since glia are intermingled with neurons in the brain and they express receptors and ion channels that are also expressed in neurons, it is difficult to perform such manipulation selectively in glial cells, while leaving neighboring neurons unaffected.
Optogenetics and chemogenetics are novel manipulation techniques based on genetically encoded effector molecules, such as specific ion channels and GPCRs, that respond to exogenously delivered light stimuli or synthetic ligands, but are unresponsive to endogenous molecules. In combination with cell type-specific promoters and other genetic tools, expression of effector molecules can be restricted to specific cell types. In addition, a controlled delivery of exogenous stimuli permits temporal precision of manipulation. Thus, the features of spatial and temporal control make it possible to perform a time-resolved functional manipulation in a specific population of cells. Optogenetics and chemogenetics have been used most extensively in the study of neuronal circuits and behavior, but they have also been employed in a number of studies focusing on glial cells, mainly astrocytes. Such studies have demonstrated that astrocytes play a critical role not only in a variety of basic physiological responses, including visual processing, norepinephrine release, breathing, cerebral blood flow, feeding, memory, and sleep, but also in pathological conditions, including drug addition, depression, and ischemia. Those studies have further revealed the importance of gliotransmitters, such as extracellular glutamate, ATP, and L-lactate, that modulate excitability and synaptic transmission in neighboring neurons. However, it is still debatable whether astrocytic release of gliotransmitters is a calcium-dependent process. In addition, the exact molecular mechanisms governing gliotransmitter release from astrocytes remains to be revealed. A combinatorial approach of advanced functional manipulation techniques such as optogenetics and chemogenetics, together with pharmacological and molecular genetic methods, can further our understanding of glial function in health and disease, including neurodevelopment, neurodegenerative disorders, and neuroinflammatory conditions.
Figures
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
{kind=link}
Tables
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
{kind=link}
| Effector | Expression | Promoter | Genetic tool | Delivery | Photostimulation | Released gliotransmitter | Response (nearby neurons) | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Response (opsin+ cells) | Response (animals) | ||||||||
| CatCh or ChR2(H134R) | Rat cortical astrocytes | GFAP | None | Txn. | Inward current, [Ca2+]i↑ | n.d. | Glutamate | sEPSC, [Ca2+]i↑ | Ref. 97 |
| CatCh or LiGluR | Mouse neocortical astrocytes | CAG | None | Txn. | [Ca2+]i↑ | n.d. | Glutamate | n.d. | Ref. 98 |
| CMV | |||||||||
| ChR2 | GL261 cell line (murine astrocytes) | CMV | None | Txn. | Na+ & Ca2+ influx, [H+]i↑ | n.d. | Glutamate | [Ca2+]i↑, apoptosis | Ref. 106 |
| ChR2, ChR2(H134R), CatCh, optoα1AR or optoβ2AR | Rat cortical astrocytes | GFAP | None | AAV | [Ca2+]i↑ | n.d. | ATP (autocrine) | n.d. | Ref. 107 |
| ChR2(C128S) | Cerebellar slices | Mlc1 | tTA/tetO | TG mice | Inward current, c-fos ↑ | n.d. | n.d. | n.d. | Ref. 110 |
| ChR2(C128S) | Cerebellar slices | Mlc1 | tTA/tetO | TG mice | Inward current | Glutamate | Inward current in PC | Ref. 111 | |
| Cerebellum ( | Mlc1 | tTA/tetO | TG mice | n.d. | Perturbation of motor behavior, pupil dilation ↑ | n.d. | c-fos in PC, SC/BC ↑ | ||
| ChR2(C128S) | Cerebellar slices | Mlc1 | tTA/tetO | TG mice | Inward current, [H+]i↑ | n.d. | Glutamate | Inward current in PC | Ref. 70 |
| ArchT | Cerebellar slices | Mlc1 | tTA/tetO | TG mice | Outward current, [H+]i↓ | n.d. | Ischemia-induced current in PC ↓ | Ref. 70 | |
| Cerebellum ( | Mlc1 | tTA/tetO | TG mice | Ischemic infarction ↓ | |||||
| ChR2(C128S) | Somatomotor cortex ( | Mlc1 | tTA/tetO | TG mice | n.d. | CBF ↑ | K+ | n.d. | Ref. 112 |
| ChR2(C128S) | V1 cortical slices, V1 ( | GFAP | None | AAV | [Ca2+]i↑ | n.d. | Glutamate | mEPSC ↑ (PV+ neuron) mixed resp. (SOM+, excitatory neuron) | Ref. 117 |
| ChR2 | Brainstem slices, primary culture | GFAP | None | AAV | [Ca2+]i↑ | ATP | Depolarization of RTN neurons | Ref. 16 | |
| VS ( | GFAP | None | AAV | n.d. | Respiratory activity ↑ | ATP | n.d. | ||
| ChR2(H134R) or optoβ2AR | Brainstem slices | GFAP | None | AAV | [H+]i↑ | n.d. | L-lactate | Depolarization of LC neurons, NE release↑ | Ref. 120 |
| ChR2 | STN ( | GFAP | None | lentivirus | n.d. | n.d. | n.d. | Firing of STN neurons ↓ | Ref. 124 |
| ChR2(H134R) | Hypothalamus ( | GFAP | None | AAV | n.d. | NREM, REM sleep ↑ | n.d. | n.d. | Ref. 125 |
| ChR2(H134R) | ACC ( | GFAP | Cre | AAV | n.d. | Wakefulnees ↑, NREM sleep ↓ | n.d. | n.d. | Ref. 126 |
| ArchT | V1 cortical slices | GFAP | Cre | AAV | [Ca2+]i↑ | Glutamate | Slow oscillation state in V1 neurons ↑, firing rate of V1 neurons ↑ | Ref. 127 | |
| V1 ( | |||||||||
AAV, adeno-associated virus; ACC, anterior cingulate cortex; ArchT, light-gated outward proton pump; BC, basket cells; ChR2, channelrhodopsin-2; CatCh, calcium translocating channelrhodopsin; CBF, cerebral blood flow; CMV, cytomegalovirus; sEPSC, spontaneous excitatory synaptic transmission; GFAP, glial acidic fibrillary acidic protein; GC, granule cell; GL261, murine astrocyte cell lines; LC, locus coeruleus; LiGluR, light-gated ionotropic glutamate receptor 6; mEPSC, miniature excitatory synaptic transmission; Mlc1, megalencephalic leukoencephalopathy with subcortical cysts 1; NE, norepinephrine; NREM, non-rapid eye movement; optoα1AR, a chimeric receptor combined rhodopsin with α1-adrenoreceptor; optoβ2AR, a chimeric receptor combined rhodopsin with β2-adrenoreceptor; PC, purkinje cells; PV, parvalbumin; REM, rapid eye movement; Resp, response; RTN, retrotrapezoid nucleus; SC, stellate cells; SOM, somatostatin; STN, subthalamic nucleus; tTA, tetracycline-controlled transcriptional activator; tetO, tTA-responsive promoter sequence; TG, transgenic; Txn, transfection; V1, primary visual cortex; VS, ventral surface of the brainstem; n.d., not determined.
{kind=link}
| Effector | Expression | Promoter | Genetic tool | Delivery | Stimulation | Released gliotransmitter | Response (nearby neurons) | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Response (effector+ cells) | Response (animals) | ||||||||
| MrgprA1 | Hippocampal slices | GFAP | tTA/tetO | TG mice | [Ca2+]i↑ | n.d. | n.d. | No response in CA1 pyramidal neurons | Ref. 42 |
| MrgprA1 | Hippocampal slices | GFAP | tTA/tetO | TG mice | sEPSC, [Ca2+]i↑ | n.d. | n.d. | No response in CA1 pyramidal neurons | Ref. 40 |
| hM3Dq | V1 ( | GFAP | Cre | AAV | [Ca2+]i↑ | No change in CBF | n.d. | n.d. | Ref. 41 |
| MrgprA1 | Primary astrocytes | GFAP | tTA/tetO | TG mice | [Ca2+]i↑ | ATP | n.d. | Ref. 138 | |
| Right cerebral ventricle, mPFC, hippocampus ( | GFAP | tTA/tetO | TG mice | ATP | |||||
| Ro1 | Whole animal (GFAP+ cells) | GFAP | tTA/tetO | TG mice | pERK↑ | Hydrocephalus | n.d. | n.d. | Ref. 139 |
| hM3Dq | Hippocampal slices | GFAP | None | TG mice | [Ca2+]i↑ | ||||
| Whole animal (GFAP+ cells) | GFAP | None | TG mice | Change in ANS-mediated responses and motor behaviors | n.d. | n.d. | Ref. 142 | ||
| hM3Dq | Intestine | GFAP | tTA/tetO | TG mice | [Ca2+]i↑ | Gut contraction ↑ | |||
| hM3Dq | Whole animal | GFAP | creERT2 | TG mice | n.d. | Hypothermia | n.d. | n.d. | Ref. 143 |
| Rs1 | Whole animal, primary astrocytes | GFAP | tTA/tetO | TG mice, lentivirus | cAMP↑, pERK↑, pCREB↑ | Memory function ↓ | n.d. | n.d. | Ref. 144 |
| hM3Dq | Arcuate nucleus ( | GFAP | None | AAV | c-fos↑ | Ghrelin-evoked feeding ↓ | Adenosine | ||
| Hypothalamic slices | GFAP | None | AAV | n.d. | Adenosine | n.d. | Ref. 147 | ||
| hM4Di | Arcuate nucleus ( | GFAP | None | AAV | no change in c-fos | Ghrelin-evoked feeding ↑ | n.d. | n.d. | Ref. 147 |
| hM3Dq | Primary astrocytes | GFAP | None | AAV | [Ca2+]i↑ | n.d. | n.d. | Ref. 154 | |
| NAcore ( | GFAP | None | AAV | [Ca2+]i↑ | Ethanol seeking ↓ | ||||
| hM3Dq | NAcore ( | GFAP | None | AAV | n.d. | Cue-induced reinstatement of cocaine seeking ↓ | Glutamate | n.d. | Ref. 155 |
AAV, adeno-associated virus; ANS, autonomic nervous system; AgRP, agouti-related protein; CA1, cornu ammonis area 1; CBF, cerebral blood flow; cAMP, cyclic adenosine monophosphate; pCREB, phosphorylated cAMP response element binding protein; creERT2, tamoxifen-inducible cre; sEPSC, spontaneous excitatory synaptic transmission; pERK, phosphorylated extracellular signal-regulated kinase; GFAP, glial acidic fibrillary acidic protein; mPFC, medial prefrontal cortex; MrgprA1, mas-related G protein-coupled receptor A1; NAcore, nucleus accumbens core; tTA, tetracycline-controlled transcriptional activator; tetO, tTA-responsive promoter sequence; TG, transgenic; V1, primary visual cortex; n.d., not determined.
References
- Go to
- Abstract
- INTRODUCTION
- OPTOGENETICS AND CHEMOGENETICS
- OPTOGENETIC STIMULATION OF CULTURED ASTROCYTES
- OPTOGENETIC STIMULATION OF ASTROCYTES USING TRANSGENIC MICE
- OPTOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED OPSINS
- CHEMOGENETIC STIMULATION OF ASTROCYTES IN TRANSGENIC MICE
- CHEMOGENETIC STIMULATION OF ASTROCYTES EXPRESSING VIRALLY DELIVERED EFFECTORS
- CONCLUDING REMARKS
- Figure
- Table
- Reference
- Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci 1999;22:208-215.

- Halassa MM, Fellin T, Haydon PG. The tripartite synapse: roles for gliotransmission in health and disease. Trends Mol Med 2007;13:54-63.
- Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci 2009;32:421-431.
- Ventura R, Harris KM. Three-dimensional relationships between hippocampal synapses and astrocytes. J Neurosci 1999;19:6897-6906.
- Danbolt NC. Glutamate uptake. Prog Neurobiol 2001;65:1-105.
- Huang YH, Bergles DE. Glutamate transporters bring competition to the synapse. Curr Opin Neurobiol 2004;14:346-352.
- Rothstein JD, Martin L, Levey AI, Dykes-Hoberg M, Jin L, Wu D, Nash N, Kuncl RW. Localization of neuronal and glial glutamate transporters. Neuron 1994;13:713-725.
- Sutherland ML, Delaney TA, Noebels JL. Glutamate transporter mRNA expression in proliferative zones of the developing and adult murine CNS. J Neurosci 1996;16:2191-2207.
- Tzingounis AV, Wadiche JI. Glutamate transporters: confining runaway excitation by shaping synaptic transmission. Nat Rev Neurosci 2007;8:935-947.
- Dirnagl U, Iadecola C, Moskowitz MA. Pathobiology of ischaemic stroke: an integrated view. Trends Neurosci 1999;22:391-397.
- Sloan SA, Barres BA. The detrimental role of glial acidification during ischemia. Neuron 2014;81:221-223.
- Szydlowska K, Tymianski M. Calcium, ischemia and excitotoxicity. Cell Calcium 2010;47:122-129.
- Araque A, Carmignoto G, Haydon PG. Dynamic signaling between astrocytes and neurons. Annu Rev Physiol 2001;63:795-813.
- Bezzi P, Volterra A. A neuron-glia signalling network in the active brain. Curr Opin Neurobiol 2001;11:387-394.
- Gordon GR, Iremonger KJ, Kantevari S, Ellis-Davies GC, MacVicar BA, Bains JS. Astrocyte-mediated distributed plasticity at hypothalamic glutamate synapses. Neuron 2009;64:391-403.
- Gourine AV, Kasymov V, Marina N, Tang F, Figueiredo MF, Lane S, Teschemacher AG, Spyer KM, Deisseroth K, Kasparov S. Astrocytes control breathing through pH-dependent release of ATP. Science 2010;329:571-575.
- Henneberger C, Papouin T, Oliet SH, Rusakov DA. Long-term potentiation depends on release of D-serine from astrocytes. Nature 2010;463:232-236.
- Wang F, Smith NA, Xu Q, Fujita T, Baba A, Matsuda T, Takano T, Bekar L, Nedergaard M. Astrocytes modulate neural network activity by Ca2+-dependent uptake of extracellular K+. Sci Signal 2012;5:ra26.
- Agulhon C, Petravicz J, McMullen AB, Sweger EJ, Minton SK, Taves SR, Casper KB, Fiacco TA, McCarthy KD. What is the role of astrocyte calcium in neurophysiology?. Neuron 2008;59:932-946.
- Petravicz J, Fiacco TA, McCarthy KD. Loss of IP3 receptor-dependent Ca2+ increases in hippocampal astrocytes does not affect baseline CA1 pyramidal neuron synaptic activity. J Neurosci 2008;28:4967-4973.
- Sheppard CA, Simpson PB, Sharp AH, Nucifora FC, Ross CA, Lange GD, Russell JT. Comparison of type 2 inositol 1,4,5-trisphosphate receptor distribution and subcellular Ca2+ release sites that support Ca2+ waves in cultured astrocytes. J Neurochem 1997;68:2317-2327.
- Liu QS, Xu Q, Arcuino G, Kang J, Nedergaard M. Astrocyte-mediated activation of neuronal kainate receptors. Proc Natl Acad Sci U S A 2004;101:3172-3177.
- Liu QS, Xu Q, Kang J, Nedergaard M. Astrocyte activation of presynaptic metabotropic glutamate receptors modulates hippocampal inhibitory synaptic transmission. Neuron Glia Biol 2004;1:307-316.
- Sasaki T, Kuga N, Namiki S, Matsuki N, Ikegaya Y. Locally synchronized astrocytes. Cereb Cortex 2011;21:1889-1900.
- Bender AS, Norenberg MD. Effect of ammonia on GABA uptake and release in cultured astrocytes. Neurochem Int 2000;36:389-395.
- Cotrina ML, Lin JH, Alves-Rodrigues A, Liu S, Li J, Azmi-Ghadimi H, Kang J, Naus CC, Nedergaard M. Connexins regulate calcium signaling by controlling ATP release. Proc Natl Acad Sci U S A 1998;95:15735-15740.
- Duan S, Anderson CM, Keung EC, Chen Y, Chen Y, Swanson RA. P2X7 receptor-mediated release of excitatory amino acids from astrocytes. J Neurosci 2003;23:1320-1328.
- Haskew-Layton RE, Rudkouskaya A, Jin Y, Feustel PJ, Kimelberg HK, Mongin AA. Two distinct modes of hypoosmotic medium-induced release of excitatory amino acids and taurine in the rat brain in vivo. PLoS One 2008;3:e3543.
- Kimelberg HK, Goderie SK, Higman S, Pang S, Waniewski RA. Swelling-induced release of glutamate, aspartate, and taurine from astrocyte cultures. J Neurosci 1990;10:1583-1591.
- Kukley M, Barden JA, Steinhäuser C, Jabs R. Distribution of P2X receptors on astrocytes in juvenile rat hippocampus. Glia 2001;36:11-21.
- Lee S, Yoon BE, Berglund K, Oh SJ, Park H, Shin HS, Augustine GJ, Lee CJ. Channel-mediated tonic GABA release from glia. Science 2010;330:790-796.
- Malarkey EB, Parpura V. Mechanisms of glutamate release from astrocytes. Neurochem Int 2008;52:142-154.
- Mongin AA, Kimelberg HK. ATP potently modulates anion channel-mediated excitatory amino acid release from cultured astrocytes. Am J Physiol Cell Physiol 2002;283:C569-C578.
- Moran MM, Melendez R, Baker D, Kalivas PW, Seamans JK. Cystine/glutamate antiporter regulation of vesicular glutamate release. Ann N Y Acad Sci 2003;1003:445-447.
- Shanker G, Aschner M. Identification and characterization of uptake systems for cystine and cysteine in cultured astrocytes and neurons: evidence for methylmercury-targeted disruption of astrocyte transport. J Neurosci Res 2001;66:998-1002.
- Tang XC, Kalivas PW. Bidirectional modulation of cystine/glutamate exchanger activity in cultured cortical astrocytes. Ann N Y Acad Sci 2003;1003:472-475.
- Woo DH, Han KS, Shim JW, Yoon BE, Kim E, Bae JY, Oh SJ, Hwang EM, Marmorstein AD, Bae YC, Park JY, Lee CJ. TREK-1 and Best1 channels mediate fast and slow glutamate release in astrocytes upon GPCR activation. Cell 2012;151:25-40.
- Ye ZC, Wyeth MS, Baltan-Tekkok S, Ransom BR. Functional hemichannels in astrocytes: a novel mechanism of glutamate release. J Neurosci 2003;23:3588-3596.
- Stout CE, Costantin JL, Naus CC, Charles AC. Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels. J Biol Chem 2002;277:10482-10488.
- Agulhon C, Fiacco TA, McCarthy KD. Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling. Science 2010;327:1250-1254.
- Bonder DE, McCarthy KD. Astrocytic Gq-GPCR-linked IP3R-dependent Ca2+ signaling does not mediate neurovascular coupling in mouse visual cortex in vivo. J Neurosci 2014;34:13139-13150.
- Fiacco TA, Agulhon C, Taves SR, Petravicz J, Casper KB, Dong X, Chen J, McCarthy KD. Selective stimulation of astrocyte calcium in situ does not affect neuronal excitatory synaptic activity. Neuron 2007;54:611-626.
- Hamilton NB, Attwell D. Do astrocytes really exocytose neurotransmitters?. Nat Rev Neurosci 2010;11:227-238.
- Santello M, Calì C, Bezzi P. Gliotransmission and the tripartite synapse. Adv Exp Med Biol 2012;970:307-331.
- Shigetomi E, Bowser DN, Sofroniew MV, Khakh BS. Two forms of astrocyte calcium excitability have distinct effects on NMDA receptor-mediated slow inward currents in pyramidal neurons. J Neurosci 2008;28:6659-6663.
- Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci 2005;8:1263-1268.
- Tye KM, Deisseroth K. Optogenetic investigation of neural circuits underlying brain disease in animal models. Nat Rev Neurosci 2012;13:251-266.
- Bernstein JG, Boyden ES. Optogenetic tools for analyzing the neural circuits of behavior. Trends Cogn Sci 2011;15:592-600.
- Hegemann P, Nagel G. From channelrhodopsins to optogenetics. EMBO Mol Med 2013;5:173-176.
- Nagel G, Ollig D, Fuhrmann M, Kateriya S, Musti AM, Bamberg E, Hegemann P. Channelrhodopsin-1: a light-gated proton channel in green algae. Science 2002;296:2395-2398.
- Nagel G, Szellas T, Huhn W, Kateriya S, Adeishvili N, Berthold P, Ollig D, Hegemann P, Bamberg E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc Natl Acad Sci U S A 2003;100:13940-13945.
- Lin JY, Lin MZ, Steinbach P, Tsien RY. Characterization of engineered channelrhodopsin variants with improved properties and kinetics. Biophys J 2009;96:1803-1814.
- Bamann C, Gueta R, Kleinlogel S, Nagel G, Bamberg E. Structural guidance of the photocycle of channelrhodopsin-2 by an interhelical hydrogen bond. Biochemistry 2010;49:267-278.
- Berndt A, Schoenenberger P, Mattis J, Tye KM, Deisseroth K, Hegemann P, Oertner TG. High-efficiency channelrhodopsins for fast neuronal stimulation at low light levels. Proc Natl Acad Sci U S A 2011;108:7595-7600.
- Berndt A, Yizhar O, Gunaydin LA, Hegemann P, Deisseroth K. Bi-stable neural state switches. Nat Neurosci 2009;12:229-234.
- Gunaydin LA, Yizhar O, Berndt A, Sohal VS, Deisseroth K, Hegemann P. Ultrafast optogenetic control. Nat Neurosci 2010;13:387-392.
- Mattis J, Tye KM, Ferenczi EA, Ramakrishnan C, O'Shea DJ, Prakash R, Gunaydin LA, Hyun M, Fenno LE, Gradinaru V, Yizhar O, Deisseroth K. Principles for applying optogenetic tools derived from direct comparative analysis of microbial opsins. Nat Methods 2011;9:159-172.
- Kleinlogel S, Feldbauer K, Dempski RE, Fotis H, Wood PG, Bamann C, Bamberg E. Ultra light-sensitive and fast neuronal activation with the Ca2+-permeable channelrhodopsin CatCh. Nat Neurosci 2011;14:513-518.
- Prigge M, Schneider F, Tsunoda SP, Shilyansky C, Wietek J, Deisseroth K, Hegemann P. Color-tuned channelrhodopsins for multiwavelength optogenetics. J Biol Chem 2012;287:31804-31812.
- Schneider F, Gradmann D, Hegemann P. Ion selectivity and competition in channelrhodopsins. Biophys J 2013;105:91-100.
- Volgraf M, Gorostiza P, Numano R, Kramer RH, Isacoff EY, Trauner D. Allosteric control of an ionotropic glutamate receptor with an optical switch. Nat Chem Biol 2006;2:47-52.
- Szobota S, Gorostiza P, Del Bene F, Wyart C, Fortin DL, Kolstad KD, Tulyathan O, Volgraf M, Numano R, Aaron HL, Scott EK, Kramer RH, Flannery J, Baier H, Trauner D, Isacoff EY. Remote control of neuronal activity with a light-gated glutamate receptor. Neuron 2007;54:535-545.
- Gradinaru V, Thompson KR, Deisseroth K. eNpHR: a Natronomonas halorhodopsin enhanced for optogenetic applications. Brain Cell Biol 2008;36:129-139.
- Gradinaru V, Zhang F, Ramakrishnan C, Mattis J, Prakash R, Diester I, Goshen I, Thompson KR, Deisseroth K. Molecular and cellular approaches for diversifying and extending optogenetics. Cell 2010;141:154-165.
- Han X, Boyden ES. Multiple-color optical activation, silencing, and desynchronization of neural activity, with single-spike temporal resolution. PLoS One 2007;2:e299.
- Lanyi JK. Halorhodopsin, a light-driven electrogenic chloride-transport system. Physiol Rev 1990;70:319-330.
- Zhang F, Wang LP, Brauner M, Liewald JF, Kay K, Watzke N, Wood PG, Bamberg E, Nagel G, Gottschalk A, Deisseroth K. Multimodal fast optical interrogation of neural circuitry. Nature 2007;446:633-639.
- Zhao S, Cunha C, Zhang F, Liu Q, Gloss B, Deisseroth K, Augustine GJ, Feng G. Improved expression of halorhodopsin for light-induced silencing of neuronal activity. Brain Cell Biol 2008;36:141-154.
- Kolbe M, Besir H, Essen LO, Oesterhelt D. Structure of the light-driven chloride pump halorhodopsin at 1.8 A resolution. Science 2000;288:1390-1396.
- Beppu K, Sasaki T, Tanaka KF, Yamanaka A, Fukazawa Y, Shigemoto R, Matsui K. Optogenetic countering of glial acidosis suppresses glial glutamate release and ischemic brain damage. Neuron 2014;81:314-320.
- Chow BY, Han X, Dobry AS, Qian X, Chuong AS, Li M, Henninger MA, Belfort GM, Lin Y, Monahan PE, Boyden ES. High-performance genetically targetable optical neural silencing by light-driven proton pumps. Nature 2010;463:98-102.
- Mahn M, Prigge M, Ron S, Levy R, Yizhar O. Biophysical constraints of optogenetic inhibition at presynaptic terminals. Nat Neurosci 2016;19:554-556.
- Airan RD, Thompson KR, Fenno LE, Bernstein H, Deisseroth K. Temporally precise in vivo control of intracellular signalling. Nature 2009;458:1025-1029.
- Bailes HJ, Zhuang LY, Lucas RJ. Reproducible and sustained regulation of G alpha s signalling using a metazoan opsin as an optogenetic tool. PLoS One 2012;7:e30774.
- Armbruster BN, Li X, Pausch MH, Herlitze S, Roth BL. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci U S A 2007;104:5163-5168.
- Dong S, Rogan SC, Roth BL. Directed molecular evolution of DREADDs: a generic approach to creating next-generation RASSLs. Nat Protoc 2010;5:561-573.
- Nichols CD, Roth BL. Engineered G-protein coupled receptors are powerful tools to investigate biological processes and behaviors. Front Mol Neurosci 2009;2:16.
- Armbruster B, Roth B. Creation of designer biogenic amine receptors via directed molecular evolution. Neuropsychopharmacology 2005;30:S265.
- Alexander GM, Rogan SC, Abbas AI, Armbruster BN, Pei Y, Allen JA, Nonneman RJ, Hartmann J, Moy SS, Nicolelis MA, McNamara JO, Roth BL. Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 2009;63:27-39.
- Atasoy D, Betley JN, Su HH, Sternson SM. Deconstruction of a neural circuit for hunger. Nature 2012;488:172-177.
- Kong D, Tong Q, Ye C, Koda S, Fuller PM, Krashes MJ, Vong L, Ray RS, Olson DP, Lowell BB. GABAergic RIP-Cre neurons in the arcuate nucleus selectively regulate energy expenditure. Cell 2012;151:645-657.
- Krashes MJ, Koda S, Ye C, Rogan SC, Adams AC, Cusher DS, Maratos-Flier E, Roth BL, Lowell BB. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J Clin Invest 2011;121:1424-1428.
- Krashes MJ, Shah BP, Koda S, Lowell BB. Rapid versus delayed stimulation of feeding by the endogenously released AgRP neuron mediators GABA, NPY, and AgRP. Cell Metab 2013;18:588-595.
- Nawaratne V, Leach K, Suratman N, Loiacono RE, Felder CC, Armbruster BN, Roth BL, Sexton PM, Christopoulos A. New insights into the function of M4 muscarinic acetylcholine receptors gained using a novel allosteric modulator and a DREADD (designer receptor exclusively activated by a designer drug). Mol Pharmacol 2008;74:1119-1131.
- Kunkel MT, Peralta EG. Identification of domains conferring G protein regulation on inward rectifier potassium channels. Cell 1995;83:443-449.
- Reuveny E, Slesinger PA, Inglese J, Morales JM, Iñiguez-Lluhi JA, Lefkowitz RJ, Bourne HR, Jan YN, Jan LY. Activation of the cloned muscarinic potassium channel by G protein beta gamma subunits. Nature 1994;370:143-146.
- Bonni A, Sun Y, Nadal-Vicens M, Bhatt A, Frank DA, Rozovsky I, Stahl N, Yancopoulos GD, Greenberg ME. Regulation of gliogenesis in the central nervous system by the JAK-STAT signaling pathway. Science 1997;278:477-483.
- Brenner M, Kisseberth WC, Su Y, Besnard F, Messing A. GFAP promoter directs astrocyte-specific expression in transgenic mice. J Neurosci 1994;14:1030-1037.
- de Leeuw B, Su M, ter Horst M, Iwata S, Rodijk M, Hoeben RC, Messing A, Smitt PS, Brenner M. Increased gliaspecific transgene expression with glial fibrillary acidic protein promoters containing multiple enhancer elements. J Neurosci Res 2006;83:744-753.
- Dymecki SM, Ray RS, Kim JC. Mapping cell fate and function using recombinase-based intersectional strategies. Methods Enzymol 2010;477:183-213.
- Luo L, Callaway EM, Svoboda K. Genetic dissection of neural circuits. Neuron 2008;57:634-660.
- Sprengel R, Hasan MT. Tetracycline-controlled genetic switches. Handb Exp Pharmacol 2007;:49-72.
- Tanaka KF, Ahmari SE, Leonardo ED, Richardson-Jones JW, Budreck EC, Scheiffele P, Sugio S, Inamura N, Ikenaka K, Hen R. Flexible Accelerated STOP Tetracycline Operator-knockin (FAST): a versatile and efficient new gene modulating system. Biol Psychiatry 2010;67:770-773.
- Bender D, Holschbach M, Stöcklin G. Synthesis of n.c.a. carbon-11 labelled clozapine and its major metabolite clozapine-N-oxide and comparison of their biodistribution in mice. Nucl Med Biol 1994;21:921-925.
- Jain S, Ruiz de Azua I, Lu H, White MF, Guettier JM, Wess J. Chronic activation of a designer Gq-coupled receptor improves β cell function. J Clin Invest 2013;123:1750-1762.
- Wess J, Nakajima K, Jain S. Novel designer receptors to probe GPCR signaling and physiology. Trends Pharmacol Sci 2013;34:385-392.
- Berlinguer-Palmini R, Narducci R, Merhan K, Dilaghi A, Moroni F, Masi A, Scartabelli T, Landucci E, Sili M, Schettini A, McGovern B, Maskaant P, Degenaar P, Mannaioni G. Arrays of microLEDs and astrocytes: biological amplifiers to optogenetically modulate neuronal networks reducing light requirement. PLoS One 2014;9:e108689.
- Li D, Hérault K, Isacoff EY, Oheim M, Ropert N. Optogenetic activation of LiGluR-expressing astrocytes evokes anion channel-mediated glutamate release. J Physiol 2012;590:855-873.
- Angulo MC, Kozlov AS, Charpak S, Audinat E. Glutamate released from glial cells synchronizes neuronal activity in the hippocampus. J Neurosci 2004;24:6920-6927.
- Bowman EJ, Siebers A, Altendorf K. Bafilomycins: a class of inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells. Proc Natl Acad Sci U S A 1988;85:7972-7976.
- Park H, Oh SJ, Han KS, Woo DH, Park H, Mannaioni G, Traynelis SF, Lee CJ. Bestrophin-1 encodes for the Ca2+-activated anion channel in hippocampal astrocytes. J Neurosci 2009;29:13063-13073.
- Pasti L, Zonta M, Pozzan T, Vicini S, Carmignoto G. Cytosolic calcium oscillations in astrocytes may regulate exocytotic release of glutamate. J Neurosci 2001;21:477-484.
- Zhou Q, Petersen CC, Nicoll RA. Effects of reduced vesicular filling on synaptic transmission in rat hippocampal neurones. J Physiol 2000;525:195-206.
- Araque A, Li N, Doyle RT, Haydon PG. SNARE protein-dependent glutamate release from astrocytes. J Neurosci 2000;20:666-673.
- Takano T, Kang J, Jaiswal JK, Simon SM, Lin JH, Yu Y, Li Y, Yang J, Dienel G, Zielke HR, Nedergaard M. Receptor-mediated glutamate release from volume sensitive channels in astrocytes. Proc Natl Acad Sci U S A 2005;102:16466-16471.
- Ono K, Suzuki H, Higa M, Tabata K, Sawada M. Glutamate release from astrocyte cell-line GL261 via alterations in the intracellular ion environment. J Neural Transm (Vienna) 2014;121:245-257.
- Figueiredo M, Lane S, Stout RF, Liu B, Parpura V, Teschemacher AG, Kasparov S. Comparative analysis of optogenetic actuators in cultured astrocytes. Cell Calcium 2014;56:208-214.
- Wang TF, Guidotti G. CD39 is an ecto-(Ca2+, Mg2+)-apyrase. J Biol Chem 1996;271:9898-9901.
- Gossen M, Bujard H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc Natl Acad Sci U S A 1992;89:5547-5551.
- Tanaka KF, Matsui K, Sasaki T, Sano H, Sugio S, Fan K, Hen R, Nakai J, Yanagawa Y, Hasuwa H, Okabe M, Deisseroth K, Ikenaka K, Yamanaka A. Expanding the repertoire of optogenetically targeted cells with an enhanced gene expression system. Cell Rep 2012;2:397-406.
- Sasaki T, Beppu K, Tanaka KF, Fukazawa Y, Shigemoto R, Matsui K. Application of an optogenetic byway for perturbing neuronal activity via glial photostimulation. Proc Natl Acad Sci U S A 2012;109:20720-20725.
- Masamoto K, Unekawa M, Watanabe T, Toriumi H, Takuwa H, Kawaguchi H, Kanno I, Matsui K, Tanaka KF, Tomita Y, Suzuki N. Unveiling astrocytic control of cerebral blood flow with optogenetics. Sci Rep 2015;5:11455.
- Chrissobolis S, Ziogas J, Chu Y, Faraci FM, Sobey CG. Role of inwardly rectifying K+ channels in K+-induced cerebral vasodilatation in vivo. Am J Physiol Heart Circ Physiol 2000;279:H2704-H2712.
- DiFrancesco D, Ferroni A, Visentin S. Barium-induced blockade of the inward rectifier in calf Purkinje fibres. Pflugers Arch 1984;402:446-453.
- Hirano Y, Hiraoka M. Barium-induced automatic activity in isolated ventricular myocytes from guinea-pig hearts. J Physiol 1988;395:455-472.
- An H, Cho DW, Lee SE, Yang YS, Han SC, Lee CJ. Differential cellular tropism of lentivirus and adeno-associated virus in the brain of cynomolgus monkey. Exp Neurobiol 2016;25:48-54.
- Perea G, Yang A, Boyden ES, Sur M. Optogenetic astrocyte activation modulates response selectivity of visual cortex neurons in vivo. Nat Commun 2014;5:3262.
- Feldman JL, Mitchell GS, Nattie EE. Breathing: rhythmicity, plasticity, chemosensitivity. Annu Rev Neurosci 2003;26:239-266.
- Mulkey DK, Stornetta RL, Weston MC, Simmons JR, Parker A, Bayliss DA, Guyenet PG. Respiratory control by ventral surface chemoreceptor neurons in rats. Nat Neurosci 2004;7:1360-1369.
- Tang F, Lane S, Korsak A, Paton JF, Gourine AV, Kasparov S, Teschemacher AG. Lactate-mediated glia-neuronal signalling in the mammalian brain. Nat Commun 2014;5:3284.
- Gibbs ME, Hertz L. Inhibition of astrocytic energy metabolism by D-lactate exposure impairs memory. Neurochem Int 2008;52:1012-1018.
- Newman LA, Korol DL, Gold PE. Lactate produced by glycogenolysis in astrocytes regulates memory processing. PLoS One 2011;6:e28427.
- Suzuki A, Stern SA, Bozdagi O, Huntley GW, Walker RH, Magistretti PJ, Alberini CM. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011;144:810-823.
- Gradinaru V, Mogri M, Thompson KR, Henderson JM, Deisseroth K. Optical deconstruction of parkinsonian neural circuitry. Science 2009;324:354-359.
- Pelluru D, Konadhode RR, Bhat NR, Shiromani PJ. Optogenetic stimulation of astrocytes in the posterior hypothalamus increases sleep at night in C57BL/6J mice. Eur J Neurosci 2016;43:1298-1306.
- Yamashita A, Hamada A, Suhara Y, Kawabe R, Yanase M, Kuzumaki N, Narita M, Matsui R, Okano H, Narita M. Astrocytic activation in the anterior cingulate cortex is critical for sleep disorder under neuropathic pain. Synapse 2014;68:235-247.
- Poskanzer KE, Yuste R. Astrocytes regulate cortical state switching in vivo. Proc Natl Acad Sci U S A 2016;113:E2675-E2684.
- Dong X, Han S, Zylka MJ, Simon MI, Anderson DJ. A diverse family of GPCRs expressed in specific subsets of nociceptive sensory neurons. Cell 2001;106:619-632.
- Han SK, Dong X, Hwang JI, Zylka MJ, Anderson DJ, Simon MI. Orphan G protein-coupled receptors MrgA1 and MrgC11 are distinctively activated by RF-amide-related peptides through the Gαq/11 pathway. Proc Natl Acad Sci U S A 2002;99:14740-14745.
- Andersson M, Hanse E. Astrocytes impose postburst depression of release probability at hippocampal glutamate synapses. J Neurosci 2010;30:5776-5780.
- Gómez-Gonzalo M, Losi G, Chiavegato A, Zonta M, Cammarota M, Brondi M, Vetri F, Uva L, Pozzan T, de Curtis M, Ratto GM, Carmignoto G. An excitatory loop with astrocytes contributes to drive neurons to seizure threshold. PLoS Biol 2010;8:e1000352.
- Halassa MM, Haydon PG. Integrated brain circuits: astrocytic networks modulate neuronal activity and behavior. Annu Rev Physiol 2010;72:335-355.
- Hertle DN, Yeckel MF. Distribution of inositol-1,4,5-trisphosphate receptor isotypes and ryanodine receptor isotypes during maturation of the rat hippocampus. Neuroscience 2007;150:625-638.
- Holtzclaw LA, Pandhit S, Bare DJ, Mignery GA, Russell JT. Astrocytes in adult rat brain express type 2 inositol 1,4,5-trisphosphate receptors. Glia 2002;39:69-84.
- Perea G, Araque A. Astrocytes potentiate transmitter release at single hippocampal synapses. Science 2007;317:1083-1086.
- Sharp AH, Nucifora FC, Blondel O, Sheppard CA, Zhang C, Snyder SH, Russell JT, Ryugo DK, Ross CA. Differential cellular expression of isoforms of inositol 1,4,5-triphosphate receptors in neurons and glia in brain. J Comp Neurol 1999;406:207-220.
- Li X, Zima AV, Sheikh F, Blatter LA, Chen J. Endothelin-1-induced arrhythmogenic Ca2+ signaling is abolished in atrial myocytes of inositol-1,4,5-trisphosphate(IP3)-receptor type 2-deficient mice. Circ Res 2005;96:1274-1281.
- Cao X, Li LP, Wang Q, Wu Q, Hu HH, Zhang M, Fang YY, Zhang J, Li SJ, Xiong WC, Yan HC, Gao YB, Liu JH, Li XW, Sun LR, Zeng YN, Zhu XH, Gao TM. Astrocyte-derived ATP modulates depressive-like behaviors. Nat Med 2013;19:773-777.
- Sweger EJ, Casper KB, Scearce-Levie K, Conklin BR, McCarthy KD. Development of hydrocephalus in mice expressing the Gi-coupled GPCR Ro1 RASSL receptor in astrocytes. J Neurosci 2007;27:2309-2317.
- Coward P, Wada HG, Falk MS, Chan SD, Meng F, Akil H, Conklin BR. Controlling signaling with a specifically designed Gi-coupled receptor. Proc Natl Acad Sci U S A 1998;95:352-357.
- Redfern CH, Coward P, Degtyarev MY, Lee EK, Kwa AT, Hennighausen L, Bujard H, Fishman GI, Conklin BR. Conditional expression and signaling of a specifically designed Gi-coupled receptor in transgenic mice. Nat Biotechnol 1999;17:165-169.
- Agulhon C, Boyt KM, Xie AX, Friocourt F, Roth BL, McCarthy KD. Modulation of the autonomic nervous system and behaviour by acute glial cell Gq protein-coupled receptor activation in vivo. J Physiol 2013;591:5599-5609.
- McClain JL, Fried DE, Gulbransen BD. Agonist-evoked Ca2+ signaling in enteric glia drives neural programs that regulate intestinal motility in mice. Cell Mol Gastroenterol Hepatol 2015;1:631-645.
- Sciolino NR, Plummer NW, Chen YW, Alexander GM, Robertson SD, Dudek SM, McElligott ZA, Jensen P. Recombinase-dependent mouse lines for chemogenetic activation of genetically defined cell types. Cell Rep 2016;15:2563-2573.
- Orr AG, Hsiao EC, Wang MM, Ho K, Kim DH, Wang X, Guo W, Kang J, Yu GQ, Adame A, Devidze N, Dubal DB, Masliah E, Conklin BR, Mucke L. Astrocytic adenosine receptor A2A and Gs-coupled signaling regulate memory. Nat Neurosci 2015;18:423-434.
- Chang WC, Ng JK, Nguyen T, Pellissier L, Claeysen S, Hsiao EC, Conklin BR. Modifying ligand-induced and constitutive signaling of the human 5-HT4 receptor. PLoS One 2007;2:e1317.
- Yang L, Qi Y, Yang Y. Astrocytes control food intake by inhibiting AGRP neuron activity via adenosine A1 receptors. Cell Rep 2015;11:798-807.
- Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, Matsukura S. A role for ghrelin in the central regulation of feeding. Nature 2001;409:194-198.
- Tschöp M, Smiley DL, Heiman ML. Ghrelin induces adiposity in rodents. Nature 2000;407:908-913.
- Campfield LA, Smith FJ, Guisez Y, Devos R, Burn P. Recombinant mouse OB protein: evidence for a peripheral signal linking adiposity and central neural networks. Science 1995;269:546-549.
- Halaas JL, Boozer C, Blair-West J, Fidahusein N, Denton DA, Friedman JM. Physiological response to long-term peripheral and central leptin infusion in lean and obese mice. Proc Natl Acad Sci U S A 1997;94:8878-8883.
- Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, Rabinowitz D, Lallone RL, Burley SK, Friedman JM. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995;269:543-546.
- Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T, Collins F. Effects of the obese gene product on body weight regulation in ob/ob mice. Science 1995;269:540-543.
- Bull C, Freitas KC, Zou S, Poland RS, Syed WA, Urban DJ, Minter SC, Shelton KL, Hauser KF, Negus SS, Knapp PE, Bowers MS. Rat nucleus accumbens core astrocytes modulate reward and the motivation to self-administer ethanol after abstinence. Neuropsychopharmacology 2014;39:2835-2845.
- Scofield MD, Boger HA, Smith RJ, Li H, Haydon PG, Kalivas PW. Gq-DREADD Selectively initiates glial glutamate release and inhibits cue-induced cocaine seeking. Biol Psychiatry 2015;78:441-451.