Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Original Article
Exp Neurobiol 2017; 26(2): 82-89
Published online April 30, 2017
https://doi.org/10.5607/en.2017.26.2.82
© The Korean Society for Brain and Neural Sciences
Immunohistochemical Localization of Translationally Controlled Tumor Protein in Axon Terminals of Mouse Hippocampal Neurons
Seong-Yeon Bae1†, Vadim Sheverdin1†, Jeehye Maeng1, In Kyoon Lyoo2, Pyung-Lim Han3* and Kyunglim Lee1*
1Graduate School of Pharmaceutical Sciences, College of Pharmacy, 2Ewha Brain Institute, Department of Brain and Cognitive Sciences, College of Pharmacy, Graduate School of Pharmaceutical Sciences, 3Department of Brain and Cognitive Sciences, Brain Disease Research Institute, and Department of Chemistry and Nano Science, Ewha Womans University, Seoul 03760, Korea
Correspondence to: *To whom correspondence should be addressed.
Kyunglim Lee, TEL: 82-2-3277-3024, FAX: 82-2-3277-2851
e-mail: klyoon@ewha.ac.kr
Pyung-Lim Han, TEL: 82-2-3277-4130, FAX: 82-2-3277-3419
e-mail: plhan@ewha.ac.kr
†These authors contributed equally to this work.
Translationally controlled tumor protein (TCTP) is a cytosolic protein with microtubule stabilization and calcium-binding activities. TCTP is expressed in most organs including the nervous system. However, detailed distribution and functional significance of TCTP in the brain remain unexplored. In this study, we investigated the global and subcellular distributions of TCTP in the mouse brain. Immunohistochemical analyses with anti-TCTP revealed that TCTP was widely distributed in almost all regions of the brain including the cerebral cortex, thalamus, hypothalamus, hippocampus, and amygdala, wherein it was localized in axon tracts and axon terminals. In the hippocampus, TCTP was prominently localized to axon terminals of the perforant path in the dentate gyrus, the mossy fibers in the cornu ammonis (CA)3 region, and the Schaffer collaterals in the CA1 field, but not in cell bodies of granule cells and pyramidal neurons, and in their dendritic processes. Widespread distribution of TCTP in axon tracts and axon terminals throughout the brain suggests that TCTP is likely involved in neurotransmitter release and/or maintaining synaptic structures in the brain, and that it might have a role in maintaining synaptic functions and synaptic configurations important for normal cognitive, stress and emotional functions.
Keywords: Cognition, Immunohistochemistry, Mossy fiber, Mouse hippocampus, Translationally Controlled Tumor Protein (TCTP)
INTRODUCTION
TCTP, also known as histamine releasing factor (HRF), is a highly conserved multifunctional protein among eukaryotes across animal and plant kingdoms. In mammalian cells, TCTP is known to play a role in cell division, regeneration, microtubule stabilization, apoptosis, carcinogenesis, tumor reversion, and ion homeostasis [1]. In rodents, TCTP was expressed during neural differentiation of embryonic stem cells [2], and in the cultured hippocampal neurons [3]. In the human nervous system, TCTP and its mRNA were distributed in the cerebral cortex, thalamus, caudate nucleus, and cerebellum [4,5].
TCTP was also expressed in a higher level in human glioma cells [6]. Overexpression of TCTP in glioma cell lines led to the progression of glioma [7]. TCTP/HRF immunoreactivity was higher in the cytoplasm of intact epithelial cells than that of lipofuscin-laden cells in choroid plexus from patients with paraneoplastic cerebellar degeneration, suggesting a potential role of TCTP in the cerebrospinal fluid barrier [8]. Oxidative modification of TCTP plays an important role in the neurodegeneration in amyotrophic lateral sclerosis [9]. TCTP/HRF levels were decreased in the hippocampus of schizophrenia patients [10] and in the temporal cortex, thalamus, and caudate nucleus in patients with Down's syndrome and Alzheimer's disease [5]. However, detailed distribution and functional significance of TCTP in the brain are not clearly understood.
In the present study, we investigated global and subcellular distributions of TCTP in brain sections using a previously well-characterized anti-TCTP antibody [11,12]. Our immunohistochemical analyses indicated that TCTP was distributed globally throughout the brain, where-in it was localized in axon tracts and mostly in axon terminals in the brain.
MATERIALS AND METHODS
Eight week-old male C57BL/6 mice (n=8) were purchased from Orient Bio. Inc. (Seongnam; Gyeonggi, Korea). Mice were maintained on a 12-h light/dark cycle and fed standard rodent chow and water ad libitum. All animal experiments were performed according to the guidelines provided in National Institutes of Health (Publication No. 8523: Guide for the Care and Use of Laboratory Animals), and were approved by Ewha Womans University's Institutional Animal Care and Use Committee (Approval ID: IACUC 2012-01-029).
For tissue preparation, mice were deeply anesthetized with ethyl ether, and perfused transcardially with phosphate buffered saline (PBS), followed by 10% formalin. Brains were then excised, fixed in 10% formalin for 24 h, and embedded in paraffin. Brains were coronally cut at 5~8 µm of thickness and sections were placed on glass slides, which resulted in 1~2 stained slides per tissue of each animal.
After deparaffinization and hydration, tissue sections were boiled for 15 min in sodium citrate buffer, pH 6.0 to retrieve the epitope. Endogenous peroxidase was blocked by incubation of brain sections with 1% hydrogen peroxide for 15 min. Immunostaining was performed with ImmPress Reagent kit (Vector laboratories) according to manufacturer's instructions. Following primary antibodies were used: anti-TCTP (Abcam, ab37506, 1:1000), anti-MAP2a+2b+2c (Abcam, ab68852, 1:100), anti-NMDAR2B (Abcam, ab93610, 1:1000), and anti-synapsin II (Sigma Aldrich, S2822, 1:1000). Signals reacted with primary antibodies were substituted with normal rabbit IgG (Abcam, ab27478). Secondary antibody was reacted with ABC kit and 0.05% DAB solution (Sigma-Aldrich) was used as a chromogen. After staining with cell nuclei using Harris hematoxylin (Sigma-Aldrich), the sections were mounted with “Permount” (Fisher Scientific). Stained sections were analyzed under the microscope (Axio Scope. A1, Zeiss), and representative photographs were made with a digital camera.
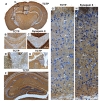
To understand the localization of TCTP in the brain, we performed immunohistochemical staining of coronal sections of mouse brains using anti-TCTP and anti-synapsin II antibodies, and analyzed the stained immunoreactivity by light microscopy. Immunohistochemical analyses indicated that TCTP was widely expressed in the brain with specific localizations in axons of neurons in almost all regions of the brain including the cerebral cortex, thalamus, hypothalamus, hippocampus, and amygdala (Fig. 1a). TCTP was localized in the lateral habenula whose expression was relatively higher than that in the medial habenula (Fig. 1e). TCTP immunoreactivity was observed in all subregions of the hippocampus, but not in the granule cells and pyramidal cells (Fig. 1f). TCTP was also distributed in all layers of the cerebral cortex, where its expression was localized in the inter-neuronal space, but TCTP immunoreactivity was barely detected in cell bodies of neurons and in dendritic processes (Fig. 1g), as interpreted from specific staining patterns by anti-TCTP and anti-synapsin II, and the latter was used as an axon fiber and axon terminal-labeling marker (Fig. 1h). TCTP immunoreactivity also was detected in the subplate, the area containing afferent fibers from the thalamus and various cortical regions (Fig. 1g).
Distinct subcellular localization of TCTP particularly to axon tracts and/or axon terminals broadly in the brain led us to examine its detailed distribution in hippocampal neurons, since the hippocampus is well-characterized for cell types, axonal tracts and synaptic circuits. The hippocampus, an important area for memory and learning processes, is comprised by distinctive subregions such as dentate gyrus (DG), cornu ammonis 3 (CA3) and CA1 regions, which have differential composition and molecular profiles. Based on the information flow through the hippocampal regions, we described the localization of TCTP from the DG to the CA3 to the CA1.
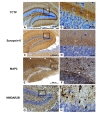
The DG is constituted with a series of strata, including polymorphic layer, stratum granulosum, and stratum moleculare. In the DG, TCTP was highly expressed in the molecular layer, but not in cell bodies of granule cells (Fig. 2a and 2b). This subcellular distribution of TCTP in the DG area was quite close to that of synapsin II (Fig. 2c and 2d). In contrast, anti-Microtubule associated protein 2 (MAP2) stained the internal side of dendritic processes of granule cells (Fig. 2e and 2f). Anti-N-methyl D-aspartate receptor subtype 2B (NMDAR2B) appeared to label the cell membrane of the dendritic processes of granule cells and cell body of astrocytes in the DG area (Fig. 2g and 2h). Thus, subcellular distributions of MAP2 and NMDAR2B were indifferent from that of TCTP. TCTP is shown to be predominantly expressed in axonal region in DG rather than in the dendrites.
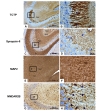
The CA regions are constituted with defined layers including stratum oriens, stratum pyramidale, stratum lucidum, stratum radiacum, stratum lacunosum, and stratum moleculare. TCTP was also found in the CA3 subregion of the hippocampus, preferentially in the stratum lucidum (Fig. 3a). At higher magnification, TCTP was found to be primarily located in axon terminals of the mossy fibers, which were originated from dentate granule cells (Fig. 3b). CA3 pyramidal cells and dendritic processes were not stained by TCTP (Fig. 3a and 3b). Anti-synapsin II staining labeled axon terminals of the mossy fibers (Fig. 3c and 3d). Anti-synapsin II staining in stratum oriens and stratum radiatum in the CA3 area was relatively higher compared to that in the stratum lucidum (Fig. 3c). MAP2 was located in the dendritic processes of CA3 pyramidal neurons (Fig. 3e and 3f ). Anti-NMDAR2B appeared to label the cell membrane of the dendritic processes of CA3 pyramidal neurons and processes of astrocytes in the CA3 area (Fig. 3g and 3h). Thus, subcellular distributions of MAP2 and NMDAR2B were not related to that of TCTP.
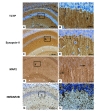
In the CA1 region, high-level TCTP immunoreactivity was found in the stratum oriens, stratum radiatum and stratum moleculare. In contrast, TCTP immunoreactivity was not detected in the stratum pyramidale which contained cell bodies of the pyramidal neurons (Fig. 4a and 4b). Dendritic processes of CA1 pyramidal cells were not stained by TCTP (Fig. 4a and 4b). To be convinced with the exact location of TCTP in CA1 subregions, we performed immunohistochemistry with additional neuronal markers. Synapsin II (axonal marker) labeled axonal terminals of neurons in most brain regions including the hippocampus, and synapsin II distribution in the CA1 was similar to that of TCTP (Fig. 4c and 4d). Anti-MAP2 (dendritic marker) stained dendrites of pyramidal cells (Fig. 4e and 4f), while anti-NMDAR2B labeled the membrane of dendritic processes (Fig. 4g and 4h). Thus, anit-MAP2 and anti-NMDAR2B staining were far different from that of TCTP or synapsin II. Taken together, these results indicate that TCTP was principally located in axon terminals rather than in dendrites and cell bodies of pyramidal cells. Because the MAP2 staining seems to be broad in CA1 regions, it is possible that TCTP is dendritically localized in other CA1 neurons.
This study is the first to characterize the detailed localization of TCTP in the hippocampus, which is well-characterized for cell types, the location of cell body and axons, and internal neural circuits. Previous studies revealed that TCTP was expressed in the human nervous tissue [13]. However, the functional significance of TCTP in brain function has been speculated on the basis of information mostly from non-neuronal cells. In the present study, we demonstrated specific localization of TCTP in axons of neurons in most brain regions including the cerebral cortex, hippocampal subregions and corpus callosum (Fig. 1).
In the CA1 region of the hippocampus, TCTP was primarily localized in axon terminals of the Schaffer collateral fibers (Fig. 4a and 4b). In the CA3 region, TCTP was preferentially localized in the stratum lucidum (Fig. 3a and 3b), the region that receives synaptic inputs of the mossy fibers and the nonmyelinated axons of the DG granule cells [14]. In the DG, TCTP was detected in the inner molecular layer (Fig. 2a and 2b), the region that receives synaptic inputs from the entorhinal cortex. In a previous study, we showed that TCTP was also highly expressed in the myenteric nerve ganglia, whereas TCTP was almost absent in perikaryon [12]. Overall, our results suggest that TCTP was predominantly localized in the axon terminals of neurons in all hippocampal subregions, than in the dendrites and cell bodies. However, the present study is primarily based on immunostaining with anti-TCTP and following light-microscopic analysis, we believe that further studies on the role of TCTP in synaptic terminals or in other subcellular regions, if any, are necessary. Also, co-localization studies using either axonal or dendritic markers are needed to define the distribution of TCTP expression in distinctive neuronal structures in the hippocampus.
The mossy fiber synapses between DG granule cells and pyramidal neurons in the CA3 region are important part of the hippocampal trisynaptic circuit [15]. The mossy fibers have large sized boutons which contain a maxi-pool of releasable vesicles and form the strong synapses with CA3 pyramidal neurons [16]. Considering that TCTP was remarkably localized in axon terminals of the mossy fibers (Fig. 3b), there is a possibility that TCTP might be involved in neurotransmitter release in mossy fibers. Given the role of microtubule stabilization and calcium-binding activities [17], we do not exclude the possibility that TCTP is involved in maintaining synaptic terminals by stabilizing cytoskeletal elements and synaptic machineries within synaptic terminals.
TCTP was shown to inhibit Na+, K+-ATPase by interacting with the 3rd cytoplasmic domain of Na+, K+-ATPase α1 subunit [18]. TCTP is also known to be involved in several diseases such as hypertension and cataract, through inhibition of Na+, K+-ATPase activity and elevating [Ca2+]i levels [19]. Of interest, the studies on the role of Na+, K+-ATPase in the synaptic transmission has been found, especially, the changes in neuronal membrane properties and the synaptic function induced by the partial inhibition of the Na+, K+-ATPase [20]. Inhibition of Na+, K+-ATPase by dihydroouabain was shown to depolarize the presynaptic terminals, which may change [Ca2+]i [21]. Therefore, it is possible that TCTP plays an role in neurotransmitter release of axons in the hippocampus, possibly through the regulation of Na+, K+-ATPase. Also, as suggested by Roque et al. in the embryonic visual system [22], TCTP possibly has a role in axonal development through the regulation of the survival signaling and axonal mitochondrial function. Further studies, including behavioral analysis using the TCTP-transgenic mouse model, might be necessary to clarify the specified roles and mechanisms of TCTP in hippocampal functions.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Bommer UA. Cellular function and regulation of the translationally controlled tumour protein TCTP. Open Allergy J 2012;5:19-32.
- Wang D, Gao L. Proteomic analysis of neural differentiation of mouse embryonic stem cells. Proteomics 2005;5:4414-4426.

- Corti V, Sanchez-Ruiz Y, Piccoli G, Bergamaschi A, Cannistraci CV, Pattini L, Cerutti S, Bachi A, Alessio M, Malgaroli A. Protein fingerprints of cultured CA3-CA1 hippocampal neurons: comparative analysis of the distribution of synaptosomal and cytosolic proteins. BMC Neurosci 2008;9:36.
- Li F, Zhang D, Fujise K. Characterization of fortilin, a novel antiapoptotic protein. J Biol Chem 2001;276:47542-47549.
- Kim SH, Cairns N, Fountoulakisc M, Lubec G. Decreased brain histamine-releasing factor protein in patients with Down syndrome and Alzheimer's disease. Neurosci Lett 2001;300:41-44.
- Gu X, Yao L, Ma G, Cui L, Li Y, Liang W, Zhao B, Li K. TCTP promotes glioma cell proliferation in vitro and in vivo via enhanced beta-catenin/TCF-4 transcription. Neuro Oncol 2014;16:217-227.
- Miao X, Chen YB, Xu SL, Zhao T, Liu JY, Li YR, Wang J, Zhang J, Guo GZ. TCTP overexpression is associated with the development and progression of glioma. Tumour Biol 2013;34:3357-3361.
- Maślińska D, Laure-Kamionowska M, Maśliński KT, Wojtecka-Łukasik E, Szukiewicz D, Maśliński S. Morphology and immuno-distribution of the histamine H4 receptor and histamine--releasing factor in choroid plexus of patients with paraneoplastic cerebellar degeneration. Inflamm Res 2009;58:45-46.
- Poon HF, Hensley K, Thongboonkerd V, Merchant ML, Lynn BC, Pierce WM, Klein JB, Calabrese V, Butterfield DA. Redox proteomics analysis of oxidatively modified proteins in G93A-SOD1 transgenic mice--a model of familial amyotrophic lateral sclerosis. Free Radic Biol Med 2005;39:453-462.
- Chung C, Tallerico T, Seeman P. Schizophrenia hippocampus has elevated expression of chondrex glycoprotein gene. Synapse 2003;50:29-34.
- Sheverdin V, Bae SY, Shin DH, Lee K. Expression and localization of translationally controlled tumor protein in rat urinary organs. Microsc Res Tech 2012;75:1576-1581.
- Sheverdin V, Jung J, Lee K. Immunohistochemical localization of translationally controlled tumor protein in the mouse digestive system. J Anat 2013;223:278-288.
- Kobayashi D, Hirayama M, Komohara Y, Mizuguchi S, Wilson Morifuji M, Ihn H, Takeya M, Kuramochi A, Araki N. Translationally controlled tumor protein is a novel biological target for neurofibromatosis type 1-associated tumors. J Biol Chem 2014;289:26314-26326.
- Gilbert PE, Brushfield AM. The role of the CA3 hippocampal subregion in spatial memory: a process oriented behavioral assessment. Prog Neuropsychopharmacol Biol Psychiatry 2009;33:774-781.
- Bischofberger J, Engel D, Frotscher M, Jonas P. Timing and efficacy of transmitter release at mossy fiber synapses in the hippocampal network. Pflugers Arch 2006;453:361-372.
- Brown TH, Johnston D. Voltage-clamp analysis of mossy fiber synaptic input to hippocampal neurons. J Neurophysiol 1983;50:487-507.
- Kim M, Jung Y, Lee K, Kim C. Identification of the calcium binding sites in translationally controlled tumor protein. Arch Pharm Res 2000;23:633-636.
- Jung J, Kim M, Kim MJ, Kim J, Moon J, Lim JS, Kim M, Lee K. Translationally controlled tumor protein interacts with the third cytoplasmic domain of Na,K-ATPase alpha subunit and inhibits the pump activity in HeLa cells. J Biol Chem 2004;279:49868-49875.
- Kim MJ, Lyu J, Sohn KB, Kim M, Cho MC, Joo CK, Lee K. Over-expression of translationally controlled tumor protein in lens epithelial cells seems to be associated with cataract development. Transgenic Res 2009;18:953-960.
- Vaillend C, Mason SE, Cuttle MF, Alger BE. Mechanisms of neuronal hyperexcitability caused by partial inhibition of Na+-K+-ATPases in the rat CA1 hippocampal region. J Neurophysiol 2002;88:2963-2978.
- Awatramani GB, Price GD, Trussell LO. Modulation of transmitter release by presynaptic resting potential and background calcium levels. Neuron 2005;48:109-121.
- Roque CG, Wong HH, Lin JQ, Holt CE. Tumor protein Tctp regulates axon development in the embryonic visual system. Development 2016;143:1134-1148.