Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Review Article
Exp Neurobiol 2022; 31(4): 221-231
Published online August 31, 2022
https://doi.org/10.5607/en22012
© The Korean Society for Brain and Neural Sciences
The Three Musketeers in the Medial Prefrontal Cortex: Subregion-specific Structural and Functional Plasticity Underlying Fear Memory Stages
Yongmin Sung and Bong-Kiun Kaang*
School of Biological Sciences, Seoul National University, Seoul 08826, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-880-7525, FAX: 82-2-884-9577
e-mail: kaang@snu.ac.kr
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Fear memory recruits various brain regions with long-lasting brain-wide subcellular events. The medial prefrontal cortex processes the emotional and cognitive functions required for adequately handling fear memory. Several studies have indicated that subdivisions within the medial prefrontal cortex, namely the prelimbic, infralimbic, and anterior cingulate cortices, may play different roles across fear memory states. Through a dedicated cytoarchitecture and connectivity, the three different regions of the medial prefrontal cortex play a specific role in maintaining and extinguishing fear memory. Furthermore, synaptic plasticity and maturation of neural circuits within the medial prefrontal cortex suggest that remote memories undergo structural and functional reorganization. Finally, recent technical advances have enabled genetic access to transiently activated neuronal ensembles within these regions, suggesting that memory trace cells in these regions may preferentially contribute to processing specific fear memory. We reviewed recently published reports and summarize the molecular, synaptic and cellular events occurring within the medial prefrontal cortex during various memory stages.
Graphical Abstract
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Keywords: Prefrontal cortex, Memory, Fear, Extinction, Neuronal plasticity
INTRODUCTION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
How is memory stored in the brain? After many years of research, several researchers have found that memory is stored in a brain-wide manner rather than in a single region. Although memory acquisition shows a high dependency on hippocampal formation, animals with hippocampal lesions experience only partial retrograde amnesia, suggesting that long-term memory recall, does not depend on the hippocampus [1]. Pavlovian fear conditioning paradigm, in which conditioned stimulus (CS) is paired with unconditioned stimulus (US) to animals, is commonly used behavioral model for the memory system. In this paradigm, fear memory ‘recall’ is a term used by researchers to describe the behavioral reactions by presenting CS to animals after initial conditioning [2]. Also, the term fear ‘extinction’ refers to the mental process underpinning a phenomenon in which animals’ fear reaction decreases following repeated exposure to CS without the US [2]. After fear memory acquisition, it is known that the short-term memory is ‘consolidated’ into the long-term memory [3]. The medial prefrontal cortex (mPFC) plays a key role in the appropriate processing of memory after consolidation. For example, human patients with deficits in the mPFC show impaired source memory, increased memory interference, and adaptation to new rules [4]. Rodent homologue of the human mPFC, including the anterior cingulate cortex (ACC), prelimbic cortex (PL) and infralimbic cortex (IL), has been well documented in the context of fear learning and memory [5-7].
It has been postulated that memory traces or engrams are stored in the form of neural ensembles [8]. Although recent memory traces have been extensively studied in the hippocampus, circuits and neural ensembles involved in long-term memory are relatively less understood. Herein, we review studies on the role of the mPFC in fear memory circuits and the role of engram cells embedded in the mPFC regions. Because the mPFC is involved in a wide range of cognitive functions, the exact role of engram cells in this region has not been well studied. However, several studies have reported that the mPFC itself plays a role in the control of innate fear [9], fear generalization [10-12] and remote fear memory [12-17] with axonal projection in amygdala or hippocampal formation. Moreover, research suggests that synaptic plasticity between engram cells underlies fear memory storage [18]. However, synaptic plasticity events occurring within the mPFC afferent and efferent regions that are correlated to memory states are yet to be summarized. In this review, we focused on recent studies reporting synaptic plasticity events within the mPFC, in the context of fear memory storage and extinction.
ANATOMICAL FEATURES OF mPFC
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Traditionally, two major criteria have been used to anatomically identify prefrontal cortex in the mammalian brain. First, cytoarchitectonic studies have revealed the laminar structure of cortical areas. In mammals, the prefrontal cortex shows an “agranular” cytoarchitecture, which is supported by a body of evidence that these areas lack cortical layer 4–specific molecular markers [19]. Although the exact parcellation of the mPFC in rodents is arguable, it is generally agreed that there are three major subdivisions of the mPFC: the prelimbic cortex (PL), infralimbic cortex (IL), and anterior cingulate cortex (ACC). These three subdivisions are distinguished by their characteristic layer structures [20]. In addition, electrophysiological investigation reports that layer II/III neurons in the PL and IL show lower excitability than those of layer V/VI, and layer II/III neurons in the PL show lower excitability than their equivalent in the IL [21].
Furthermore, early anatomical studies have found that the frontal cortex can be characterized by projections from the thalamic mediodorsal nucleus [22]. Comparison of efferent projections between the PL and IL using retrograde tracers revealed that these two regions have dramatically distinct projection patterns, suggesting their functions within the fear memory network [23-25]. In addition, the ACC, which is a part of the dorsomedial prefrontal cortex, shows innervation patterns slightly different from those of the PL and IL [26].
In addition to excitatory neurons, a heterogeneous inhibitory neuronal composition delineates the mPFC from the surrounding brain regions. Especially, IL shows relatively higher SST+ interneuron density to PV+ neuron density compared to surrounding regions including PL, in a brain-wide cell-type-specific mapping study [27]. Inhibitory interneurons consisting of approximately 17% mPFC neurons also have an intensive network within this region [28]. Monosynaptic viral tracing revealed that PL and IL share a similar long-range input pattern to SST+, PV+, and vasoactive intestinal peptide (VIP)+ interneurons [29]. While PV cells preferentially target the soma or axon initial segment of pyramidal neurons, SST cells project their axons to dendrites to inhibit dendritic Ca2+ signaling in the pyramidal neurons [30]. Interestingly, somatostatin (SST) interneurons also innervate parvalbumin (PV) interneurons within the mPFC, whereas SST+ interneurons in the visual cortex do not [31].
Recent studies have reported that these interneurons participate in learning and memory. In the PL, PV+ cells are known to be involved in fear expression [32] or memory consolidation [33], by controlling the firing patterns of pyramidal cells directly through their projection patterns. Also, SST+ interneurons in the PL were shown to have cue-specific activity after memory encoding, and that activity reappeared during fear expression, suggesting that SST+ neurons actively participate in the fear memory network in this region [34]. In addition, during working memory tasks, inhibition of SST+ cells, but not PV+ cells, impaired hippocampal–prefrontal synchrony, suggesting their role in coordinating long-range inputs from other regions [35]. In addition, one study showed that interneurons in the PL receive excitatory input from the ventral hippocampus, thereby contributing to the control of tone responsiveness of pyramidal neurons in the PL [36]. Astrocytes and microglia, which are important non-neuronal cell types in the brain, are also known to contribute to long-term memory maintenance with their dedicated memory-specific transcriptomic changes [17]. In line with this finding, one study showed that astrocytes in CA1 contribute to communication between CA1 and ACC [37]. Finally, a recent study showed that the formation of new myelination by the proliferation of oligodendrocyte precursor cells into myelinating oligodendrocytes is required for remote memory retention in the mPFC [38]. These data suggest that the local mPFC local circuit contributes to learning and memory.
RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
It is well known that the early encoding of fear memory depends on the hippocampus. However, the systems consolidation theory posits that acquired fear memory is gradually reorganized from the hippocampus to other brain structures [39]. In this context, the input from CA1 to ACC is known to mediate fear memory recall. CA1, an important hub in the memory system, has been shown to be involved in memory consolidation by a brain rhythm called a sharp-wave ripple. During sleep, when hippocampal sharp-wave ripples arise, ACC neurons fire shortly after sharp-ripple activity, but not when mice are awake [40]. Although memory-induced tight interactions during the awake state were not revealed in this study, another study showed that memory-specific interactions between these two regions arise in memory recall. Moreover, the synchronized pattern between the ACC and CA1 also indicates the age of the memory [41].
Frankland et al. [13] showed that reversible ACC inhibition with lidocaine was sufficient to impair remote fear memory but not recent memory. Goshen et al. [42] also showed that remote memory recall can be impaired by optogenetic inhibition of ACC. In support of this, another study also showed that ACC mediates generalized remote fear memory expression, by showing that ACC neural activity is required for the expression of fear response in a novel context, but not the fear-associated one [12]. After the recall of a memory, it has been postulated that memory transiently becomes “labile” and can be modified before it is reconsolidated. However, anisomycin treatment to block
Inhibition studies have suggested the role of the prelimbic and infralimbic cortices in remote fear memory. Sierra-Mercado et al. [44] showed that the vmPFC, including PL and IL, inhibition with muscimol selectively disrupted remote fear memory but not recent memory. Further studies have attempted to dissociate the roles of the prelimbic and infralimbic cortices. First, the role of the PL in fear memory was suggested by the finding that PL inactivation with tetrodotoxin during both contextual and auditory fear conditioning resulted in lowered freezing levels on the conditioning day, but not on the following day. In this study, the inactivation of PL during the retrieval session resulted in a reduced freezing level of the conditioned stimulus (CS) cue. However, PL inactivation did not affect innate fear expression, suggesting that this region plays a role in the expression of learned fear memory [45]. In addition, an early study showed that PL neurons show sustained CS response after conditioning, and their activity pattern is correlated with the fear state [46]. In summary, PL activity is necessary for remote fear memory expression, but not for recent or innate expression.
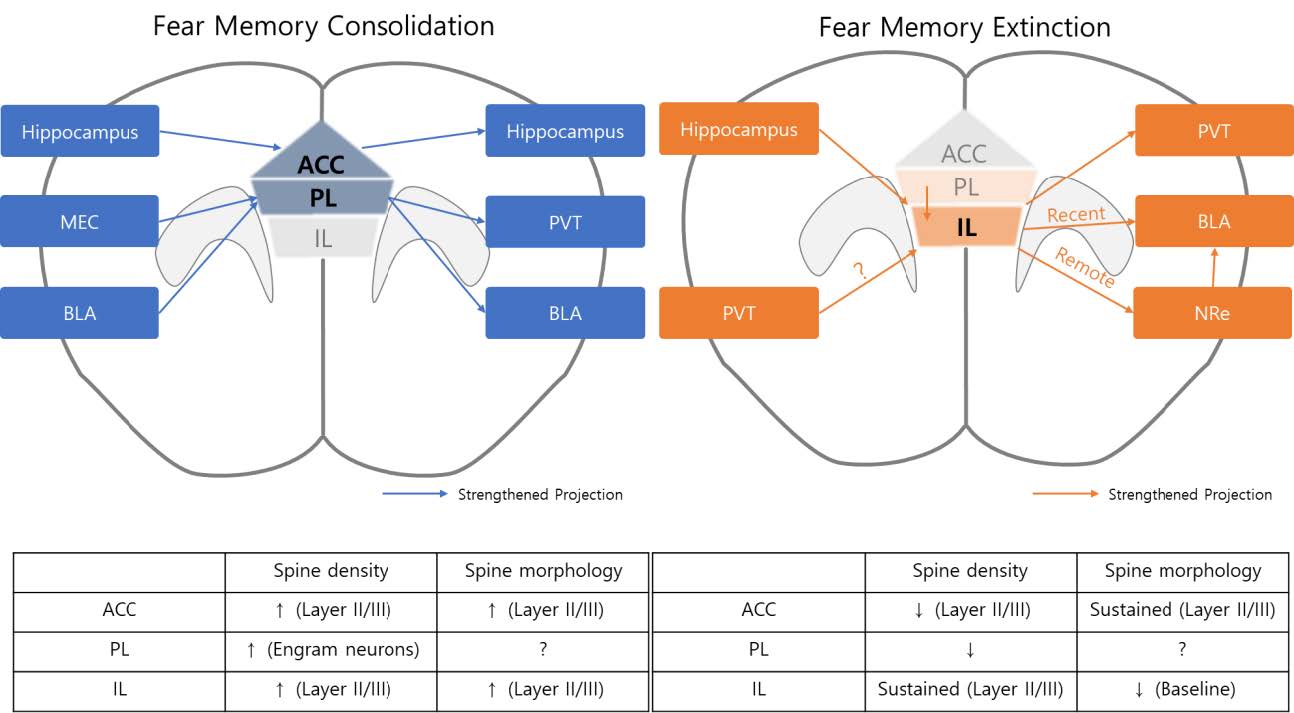
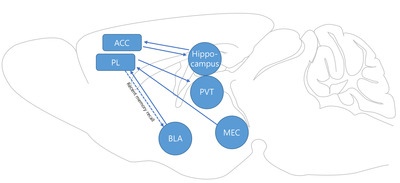
PL and basolateral amygdala (BLA) are known for encoding and expression of fear response. Although projection studies suggest that the BLA projects to both the PL and IL, differential projections toward these two regions have been observed [23]. Whereas layer II corticoamygdalar projection neurons in the PL are preferentially innervated by the BLA compared to those of the IL, layer III/V PAG (periaqueductal gray)-projecting neurons received more inputs from the BLA over those neurons of the PL, which is in agreement with the report that BLA inactivation led to a decrease in PL neuron activities [36]. In addition, maturation of fear memory recruits the connection between the PL and the PVT. Although the PL directly innervates the BLA and optogenetic inhibition of PL axons in the BLA or PL somata impaired fear recall in recent timepoints, optogenetic inhibition of the PL neurons was not sufficient to impair the fear response at remote timepoints, whereas silencing prelimbic neurons innervating the PVT impaired fear recall at remote timepoints, but not in recent ones [47]. Moreover, PVT-projecting PL neurons showed higher c-fos density than non-conditioned animals, suggesting that PL to PVT connectivity serves as a fear memory–specific circuit [47]. Laminar-specific reorganization of the prelimbic circuit involved in fear retrieval has also been reported. PL neurons projecting to the amygdala, residing in layer V, showed higher c-fos activity at 2 hours after fear conditioning, whereas layer VI PVT-projecting neurons showed higher c-fos density only at 4 days after conditioning [48]. In summary, while the ACC contributes to fear memory expression by communicating with the hippocampal formation, the PL is involved in storing and expressing remote fear memory (Fig. 1).
While the results of inhibition studies suggest that subregions of the mPFC contribute to the fear memory acquisition stage, a growing body of research suggests that neurons in these regions exhibit several types of plasticity. It is well known that temporally elevated neuronal excitability is important for neurons to be allocated and incorporated into an engram [49, 50]. Prelimbic neurons show an increased and sustained conditioned response in the conditioned tone, and higher excitability of prelimbic neurons was positively correlated with freezing behavior during fear recall and extinction failure [46]. Information from the hippocampus and amygdala, which innervate PL and fear-induced neuromodulatory inputs, is proposed to underly increase of the excitability of PL [51]. Further, the storage of fear memory in the PL requires contextual information from the MEC layer Va, because eArchT-mediated MEC layer V terminal inhibition in the PL resulted in a remote fear memory deficit, but not a recent one [52].
Notably, immediate early gene (IEG) studies also support the differential roles of the two regions. In the case of PL, studies show consistent results that fear memory recall induces fos activity in the PL. An early study using a CaMKII (Ca2+-calmodulin dependent kinase II) mutant mouse showed a gradual increase in fos induction in the ACC, PL, and IL as a function of time between fear memory acquisition and retention [13]. In addition, network analysis of brain-wide global c-fos activity showed that upregulated PL fos activity was associated with remote memory-associated fos network [53]. Similarly, a study conducted by Johannes Gräff showed higher c-fos induction in PL with remote fear recall [54].
ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Compared with studies using the fear retrieval paradigm focusing on the prelimbic cortices, reports using the extinction paradigm have shown that infralimbic cortex, which lies under the PL, participates in the extinction of fear memory. Several inhibition studies have attempted to reveal the exact role of IL in fear extinction. Inactivation of IL with muscimol and lidocaine before extinction learning was sufficient to impair the consolidation of extinction learning [55, 56]. Finally, treatment with the protein synthesis inhibitor anisomycin before extinction training affected the reconsolidation of extinction training and susceptibility to extinction training [56, 57].
With electrical microstimulation, one study showed that, in contrast to PL microstimulation which increased fear expression and decreased the effect of extinction training, rats underwent IL microstimulation showed accelerated extinction training [58]. In addition, one study revealed that IL neurons projecting to BLA are required for consolidation of fear extinction memory [59], suggesting that IL region is required for fear extinction. Optogenetic activation of IL within the context of fear conditioning also enhances effect of extinction training [60]. Furthermore, it has been shown that recall of fear extinction memory increases excitability of IL neurons temporally [61-63]. These studies suggest that the neuronal activity of IL during extinction training plays a key role in extinguishing previously learned fear response. Interestingly, it is reported that layer V/VI neurons in the IL receive excitatory input from layer V/VI neurons of the PL during extinction training [64]. In addition, synchronization of local field potential (LFP) in the fast gamma frequency between the PL and IL is enhanced during extinction [65]. Inhibition of the connection between PL and IL impaired the updating of previous association in extinction training [66]. All these results suggest that the interaction between the IL and PL might be important during extinction.
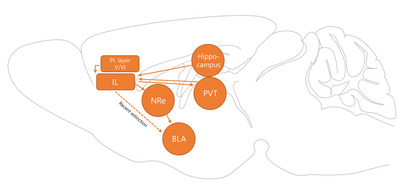
Similar to the PL, the IL circuit mediating extinction seems to undergo a time-dependent shift. Although IL itself participates in fear extinction learning at both recent and remote time points [56], a recent report showed that extinction of remote memory recruits different neurons compared to recent ones. Among the important outputs of the IL are the nucleus reuniens (NRe) and BLA. Although extinction training recruits BLA-projecting IL neurons [59], the extinction of remote fear memory did not recruit these neurons. Instead, IL neurons projecting to the NRe, which also have a strong projection to the BLA, were gradually recruited [67]. In summary, these reports show that increased activity in the IL is reflected in downstream regions that regulate extinguishing previously formed fear memory.
In addition to the PL, IL neurons also project to the PVT, and the function of this connectivity was systemically questioned by an experiment similar to that performed by Do Monte and their colleagues. When they labeled IL neurons projecting to the PVT through retrobeads, they found an increase in c-fos expression in those neurons during extinction retrieval and reported that extinction memory was not recalled when they chemogenetically inhibited IL projection in the PVT. Seven days after extinction training, optogenetic inhibition of IL projections in the PVT reversed the effect of chemogenetic activation of the PVT to the central amygdala (CeA), which lowered the freezing level during retrieval, suggesting that IL projection to the PVT mediates extinction recall [68]. PVT projection to PL is known to play a role in fear response [69]. Although PVT projects to IL more extensively than to PL, roles of axonal projection of PVT to IL in various fear memory stages are yet to be discovered. In summary, IL orchestrates extinction of fear memory with projection to PVT and BLA, which are known to be involved in fear response of animal (Fig. 2).
SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Neuronal activity can induce several types of plasticity in the brain, and the Ca2+-dependent signal cascade from the spine is known to be a key mechanism [70]. Within the spine, NMDA (N-methyl D-aspartate) receptor activation can lead to an influx of Ca2+ ions into postsynaptic neurons and activate transcription of IEGs, which are transcription factors that mediate cellular functions [71]. Within the hippocampus, GluN2A subunit-containing NMDAR is required for fear memory formation [72]. Whereas both GluN2A and GluN2B subunits are involved in long-term potentiation (LTP) induction in the ACC, GluN2B subunit-containing NMDAR function is required for early acquisition of fear memory, as microinjection of a GluN2B subunit antagonist Ro25-6981 in the ACC impaired contextual fear memory recall in mice [73].
It is well known that protein kinase Mζ (PKMζ) contributes to long-term potentiation [74], memory persistence [75], and chronic pain, another critical function of mPFC [76, 77]. Within the mPFC, prelimbic PKMζ overexpression leads to enhancement of long-term, but not short-term, fear memory [78]. Furthermore, a recent study reported that PKC and PKMζ in the PL are necessary for long-term memory reconsolidation and persistence, respectively [79]. Similarly, it is known that PKMζ and CP-AMPAR are required for long-term synaptic enhancement in ACC [80].
Structural changes initiated by neuronal activity are structural changes in the spines. In the ACC, fear conditioning increased spine density in layer II/III pyramidal neurons compared with control animals, and these effects lasted over 49 days (Table 1) [81]. A similar increase in the spine density of engram cells, which were labeled after fear conditioning, was observed in the PL (Table 1) [52]. Recently, one study showed that fear-activated CA1 engram cells had increased relative spine density connected with CA3 engram cells [18]. Unlike spine density occurring in the CA3-CA1 circuit, it is noteworthy that an increase in spine density in the ACC was not observed in the recent time window, which is consistent with inhibition studies [15]. One of the molecular mediators of spine density within the PL is a molecular motor protein (KIFs). In this study, KIF3B was upregulated 1 hour after fear conditioning, and shRNA-mediated knockdown of KIF3B led to an increase in spine density [82].
An increase in spine density suggests that prefrontal neurons are more responsive to CS cues. Simultaneously, efferent from the prefrontal cortex also undergoes strengthening, which can reinforce signals toward downstream regions underlying fear-related behavior. PL sends dense projection to paraventricular nucleus of the thalamus (PVT), and inhibition of PL-PVT connectivity at a remote time point impairs retrieval of fear memory [47].
In addition to plasticity at the dendrite level, other studies have suggested that presynaptic plasticity occurs within PFC regions. Within the ACC and infralimbic cortices, elevated GAP-43 (Growth Associated Protein 43) levels were observed, as well as c-fos [83]. Recent advances in genetics enabled genetic access of specific neurons. The Targeted Recombination in Active Populations (TRAP) system utilizes intrinsic IEG promoters to express Cre recombinase-dependent genes following neuronal activity under tamoxifen administration [84]. RNA sequencing of ACC TRAPed cells revealed that genes involved in membrane fusion are associated with long-term fear memory maintenance [17].
Long-term potentiation, where synapse function is facilitated for a long time, is known to underlie several memory functions, as well as in the engram. High-frequency stimulation results in LFP (local field potential) potentiation within the prelimbic region of rats [85]. Long-term potentiation of the hippocampus–mPFC pathway did not last for more than 1 week by HFS (high-frequency stimulation) or by exposing rats to enriched environments [86]. One interesting study showed that learning new rules was facilitated by long-term depression, showing that potentiated ventral hippocampal input to ACC carries information of learned rules, and updating internal rules requires “resetting” previous information [87].
SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
As suggested by region-level studies, it has been shown that synaptic plasticity also plays a role in extinction of acquired fear memory. By stimulating mediodorsal thalamic axons that delineate the prefrontal cortex from the surrounding cortices, Herry and Garcia [62] enabled
NMDA-dependent Ca2+ influx is important for synaptic plasticity. NMDA receptor function in the mPFC has also been studied. With viral administration of Cre recombinase and a mouse line in which Grin1 was knocked out by Cre expression, researchers achieved mPFC excitatory neuron-specific knockout (KO) of Grin1 [88]. These mPFC-NR1 KO mice showed impaired auditory fear extinction, and treatment with 3-(2-carboxypiperazin-4-yl)propyl-1-phosphonic acid (CPP), the NMDAR antagonist, into the ventral mPFC regions impaired the consolidation of extinction training [89]. In summary, NMDA-dependent synaptic plasticity events within the IL may underlie fear extinction. One of the candidate regions involved in the modification of NMDAR currents is the ventral hippocampus, which may send contextual information to the IL [90]. In a study by Soler-Cedeño et al. [90], NMDA current by the ventral hippocampus (vHP) afferents within the IL was correlated with freezing levels.
Extinction training has been shown to influence the density [91] and morphology [92] of the dendritic spine in neurons in the fear memory system. Several studies showed that extinction also induces structural changes in the mPFC regions. After extinction training, structural changes in ACC and IL neurons were reversed in a complex manner. Where extinction training affected only spine density, but not spine size, increase induced by fear acquisition in the ACC, IL neurons still showed a higher spine density than that in pseudo-conditioned animals. However, the elevation of spine size of IL neurons returned to the level observed in pseudo-conditioned mice [93]. In the PL, it is reported that apical dendrites of pyramidal neurons showed decreased spine density in animals underwent extinction training compared to no extinction group (Table 1) [94].
Recent studies suggest that several brain areas participating in remote memory systems, including the mPFC, undergo a “transition state” between the reconsolidation–extinction stage. In this study, ERK phosphorylation, which is required for fear memory reconsolidation in the amygdala [95], was observed in the mPFC during the extinction stage, whereas CREB, a downstream target of phosphorylated ERK, was observed in both the reconsolidation and extinction stages. Interestingly, between the two stages existed a transition phase, where ERK and CREB phosphorylation was downregulated to the basal level, suggesting the occurrence of “turning off” of molecular signatures in the reconsolidation stage [96].
FEAR MEMORY ENGRAM IN THE mPFC
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Many engram studies have focused on several regions that are required during memory formation, such as the hippocampal formation. PL was an early target of engram study, for inhibition studies show that inhibition of fear learning-activated neurons in this region during fear recall decreased the freezing level of mice only during remote recall [52]. As discussed earlier, spine density may explain this finding. In recent times, these neurons have not shown increased spine density, which increases with time. In addition, these neurons were reactivated during remote recall, supporting the idea that this ensemble contains actual engram cells [52]. Maturation of engram cells in the prefrontal cortex was further questioned in a recent study by the Liqun Luo group [16]. In their study, they found that 14-day-old fear memory was related to the recruitment of PL engram cell-dependent fos expression levels in cortical areas. Taking advantage of TRAPing putative engram cells, Chen et al. [17] screened ACC engram cells at a remote time point, as discussed above. By combining single-cell RNA sequencing (scRNA-seq), they screened for differentially expressed genes specific to fear memory. Although this study did not show behavioral data on whether TRAPed neurons are necessary or sufficient for fear-related behavior, genetic access to engram cells will help understand engram-specific molecular programs [17]. Finally, among other regions, IL neurons that were activated during remote recall were reactivated during extinction training, as suggested by IEG expression [97]. However, how molecular and structural properties of engram cells, compared to surrounding mPFC neurons is still missing (Table 1). Future research is needed to reveal IL engram-specific molecular changes underlying fear memory formation and extinction.
CONCLUSION
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
In this review, we summarize findings on how the mPFC contributes to fear memory. Anatomical studies and intraregional connectivity patterns show that the mPFC has a unique niche to modulate the fear response, with connection to the hippocampal area and amygdala. Surprisingly, non-neuronal cells, including oligodendrocytes, astrocytes, and microglial cells also participate in consolidating and maintaining fear memory.
Alongside these results, reports indicate that mPFC neurons show increased responsiveness to fear-related cues at remote time points compared to recent ones, indicating that slow and long-lasting changes may contribute to the consolidation of fear memory, thereby shifting fear memories from hippocampus-dependent to fewer dependent ones. Structural changes on neurons in the ACC and the PL might underlie consolidation procedure. In line with this, studies on IEG expression suggest that transiently activated neuronal ensembles or engram cells in those areas gradually mature to increase their influence on remote fear memory networks [8, 52].
Extinction training is an important mental procedure for an animal to properly adapt to the environment [2]. Behavioral results from lesion study or inhibition study suggest that the IL is required for the extinction of consolidated fear memory. Similar to the structural changes following memory consolidation, extinction training also influences structure of IL neurons [93]. Network analysis of c-fos activation pattern following extinction shows that the IL is a functional hub in the brainwide network of fear extinction [54].
Many studies indicate a functional shift of the mPFC circuit in remote memories [16, 47, 48] and extinction [67]. To date, the mechanism underlying this shift is unknown. Silent engram hypothesis posits that time-dependent maturation of engram cells makes ‘silent’ engram active [39, 52]. Structural changes in engram cells, including spine density, are reported to be altered in engram cells of the PL [52]. However, it is still unknown how structural alterations, particularly in the engram cells in three subregions of the mPFC, are linked to memory states. Future studies should reveal the molecular profiles and structural plasticity of engram cells at various memory stages.
ACKNOWLEDGEMENTS
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
This work was supported by the National Honor Scientist Program (NRF-2012R1A3A1050385) of the National Research Foundation of Korea.
Figures
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
{kind=link}
{kind=link}
Tables
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
Dendritic spine dynamics of different mPFC regions’ pyramidal neurons during different memory processes
| Region | Layer | Memory stage | Spine density | Spine morphology | Reference |
|---|---|---|---|---|---|
| ACC | II/III | Consolidation | Increased | ? | [15] |
| V/VI | Consolidation | Baseline | ? | [81] | |
| II/III | Consolidation | Increased | Enlarged | [81, 93] | |
| II/III | Extinction | Baseline | Sustained | [93] | |
| PL | - | Consolidation | Increased | ? | [52] |
| V | Extinction | Decreased | ? | [94] | |
| IL | II/III | Consolidation | Increased | Increased | [93] |
| II/III | Extinction | Sustained | Baseline | [93] |
References
- Go to
- Abstract
- Graphical Abstract
- INTRODUCTION
- ANATOMICAL FEATURES OF mPFC
- RECRUITMENT OF ACC AND PL AFTER FEAR MEMORY FORMATION
- ROLE OF THE IL IN FEAR MEMORY EXTINCTION
- SYNAPTIC PLASTICITY INDUCED BY FEAR MEMORY CONSOLIDATION
- SYNAPTIC PLASTICITY INDUCED BY FEAR EXTINCTION
- FEAR MEMORY ENGRAM IN THE mPFC
- CONCLUSION
- ACKNOWLEDGEMENTS
- Figure
- Table
- Reference
- Clark RE, Broadbent NJ, Zola SM, Squire LR (2002) Anterograde amnesia and temporally graded retrograde amnesia for a nonspatial memory task after lesions of hippocampus and subiculum. J Neurosci 22:4663-4669



- Myers KM, Davis M (2007) Mechanisms of fear extinction. Mol Psychiatry 12:120-150
- Schafe GE, Nader K, Blair HT, LeDoux JE (2001) Memory consolidation of Pavlovian fear conditioning: a cellular and molecular perspective. Trends Neurosci 24:540-546
- de Sousa AF, Chowdhury A, Silva AJ (2021) Dimensions and mechanisms of memory organization. Neuron 109:2649-2662
- Giustino TF, Maren S (2015) The role of the medial prefrontal cortex in the conditioning and extinction of fear. Front Behav Neurosci 9:298
- Marek R, Sun Y, Sah P (2019) Neural circuits for a top-down control of fear and extinction. Psychopharmacology (Berl) 236:313-320
- Dixsaut L, Gräff J (2021) The medial prefrontal cortex and fear memory: dynamics, connectivity, and engrams. Int J Mol Sci 22:12113
- Josselyn SA, Tonegawa S (2020) Memory engrams: recalling the past and imagining the future. Science 367:eaaw4325
- Jhang J, Lee H, Kang MS, Lee HS, Park H, Han JH (2018) Anterior cingulate cortex and its input to the basolateral amygdala control innate fear response. Nat Commun 9:2744
- Ortiz S, Latsko MS, Fouty JL, Dutta S, Adkins JM, Jasnow AM (2019) Anterior cingulate cortex and ventral hippocampal inputs to the basolateral amygdala selectively control generalized fear. J Neurosci 39:6526-6539
- Cullen PK, Gilman TL, Winiecki P, Riccio DC, Jasnow AM (2015) Activity of the anterior cingulate cortex and ventral hippocampus underlie increases in contextual fear generalization. Neurobiol Learn Mem 124:19-27
- Einarsson EÖ, Pors J, Nader K (2015) Systems reconsolidation reveals a selective role for the anterior cingulate cortex in generalized contextual fear memory expression. Neuropsychopharmacology 40:480-487
- Frankland PW, Bontempi B, Talton LE, Kaczmarek L, Silva AJ (2004) The involvement of the anterior cingulate cortex in remote contextual fear memory. Science 304:881-883
- Matos MR, Visser E, Kramvis I, van der Loo RJ, Gebuis T, Zalm R, Rao-Ruiz P, Mansvelder HD, Smit AB, van den Oever MC (2019) Memory strength gates the involvement of a CREB-dependent cortical fear engram in remote memory. Nat Commun 10:2315
- Restivo L, Vetere G, Bontempi B, Ammassari-Teule M (2009) The formation of recent and remote memory is associated with time-dependent formation of dendritic spines in the hippocampus and anterior cingulate cortex. J Neurosci 29:8206-8214
- DeNardo LA, Liu CD, Allen WE, Adams EL, Friedmann D, Fu L, Guenthner CJ, Tessier-Lavigne M, Luo L (2019) Temporal evolution of cortical ensembles promoting remote memory retrieval. Nat Neurosci 22:460-469
- Chen MB, Jiang X, Quake SR, Südhof TC (2020) Persistent transcriptional programmes are associated with remote memory. Nature 587:437-442
- Choi JH, Sim SE, Kim JI, Choi DI, Oh J, Ye S, Lee J, Kim T, Ko HG, Lim CS, Kaang BK (2018) Interregional synaptic maps among engram cells underlie memory formation. Science 360:430-435
- Wang X, Allen WE, Wright MA, Sylwestrak EL, Samusik N, Vesuna S, Evans K, Liu C, Ramakrishnan C, Liu J, Nolan GP, Bava FA, Deisseroth K (2018) Three-dimensional intact-tissue sequencing of single-cell transcriptional states. Science 361:eaat5691
- Van De Werd HJ, Rajkowska G, Evers P, Uylings HB (2010) Cytoarchitectonic and chemoarchitectonic characterization of the prefrontal cortical areas in the mouse. Brain Struct Funct 214:339-353
- Song C, Moyer JR Jr (2018) Layer- and subregion-specific differences in the neurophysiological properties of rat medial prefrontal cortex pyramidal neurons. J Neurophysiol 119:177-191
- Guldin WO, Pritzel M, Markowitsch HJ (1981) Prefrontal cortex of the mouse defined as cortical projection area of the thalamic mediodorsal nucleus. Brain Behav Evol 19:93-107
- Cheriyan J, Kaushik MK, Ferreira AN, Sheets PL (2016) Specific targeting of the basolateral amygdala to projectionally defined pyramidal neurons in prelimbic and infralimbic cortex. eNeuro 3:ENEURO.0002-16.2016
- Kim WB, Cho JH (2017) Synaptic targeting of double-projecting ventral CA1 hippocampal neurons to the medial prefrontal cortex and basal amygdala. J Neurosci 37:4868-4882
- Vertes RP (2004) Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse 51:32-58
- Qadir H, Krimmel SR, Mu C, Poulopoulos A, Seminowicz DA, Mathur BN (2018) Structural connectivity of the anterior cingulate cortex, claustrum, and the anterior insula of the mouse. Front Neuroanat 12:100
- Kim Y, Yang GR, Pradhan K, Venkataraju KU, Bota M, García Del Molino LC, Fitzgerald G, Ram K, He M, Levine JM, Mitra P, Huang ZJ, Wang XJ, Osten P (2017) Brain-wide maps reveal stereotyped cell-type-based cortical architecture and subcortical sexual dimorphism. Cell 171:456-469.e22
- Erö C, Gewaltig MO, Keller D, Markram H (2018) A cell atlas for the mouse brain. Front Neuroinform 12:84
- Sun Q, Li X, Ren M, Zhao M, Zhong Q, Ren Y, Luo P, Ni H, Zhang X, Zhang C, Yuan J, Li A, Luo M, Gong H, Luo Q (2019) A whole-brain map of long-range inputs to GABAergic interneurons in the mouse medial prefrontal cortex. Nat Neurosci 22:1357-1370
- Kepecs A, Fishell G (2014) Interneuron cell types are fit to function. Nature 505:318-326
- Ährlund-Richter S, Xuan Y, van Lunteren JA, Kim H, Ortiz C, Pollak Dorocic I, Meletis K, Carlén M (2019) A whole-brain atlas of monosynaptic input targeting four different cell types in the medial prefrontal cortex of the mouse. Nat Neurosci 22:657-668
- Courtin J, Chaudun F, Rozeske RR, Karalis N, Gonzalez-Campo C, Wurtz H, Abdi A, Baufreton J, Bienvenu TC, Herry C (2014) Prefrontal parvalbumin interneurons shape neuronal activity to drive fear expression. Nature 505:92-96
- Xia F, Richards BA, Tran MM, Josselyn SA, Takehara-Nishiuchi K, Frankland PW (2017) Parvalbumin-positive interneurons mediate neocortical-hippocampal interactions that are necessary for memory consolidation. Elife 6:e27868
- Cummings KA, Clem RL (2020) Prefrontal somatostatin interneurons encode fear memory. Nat Neurosci 23:61-74
- Abbas AI, Sundiang MJM, Henoch B, Morton MP, Bolkan SS, Park AJ, Harris AZ, Kellendonk C, Gordon JA (2018) Somatostatin interneurons facilitate hippocampal-prefrontal synchrony and prefrontal spatial encoding. Neuron 100:926-939.e3
- Sotres-Bayon F, Sierra-Mercado D, Pardilla-Delgado E, Quirk GJ (2012) Gating of fear in prelimbic cortex by hippocampal and amygdala inputs. Neuron 76:804-812
- Kol A, Adamsky A, Groysman M, Kreisel T, London M, Goshen I (2020) Astrocytes contribute to remote memory formation by modulating hippocampal-cortical communication during learning. Nat Neurosci 23:1229-1239
- Pan S, Mayoral SR, Choi HS, Chan JR, Kheirbek MA (2020) Preservation of a remote fear memory requires new myelin formation. Nat Neurosci 23:487-499
- Tonegawa S, Morrissey MD, Kitamura T (2018) The role of engram cells in the systems consolidation of memory. Nat Rev Neurosci 19:485-498
- Wang DV, Ikemoto S (2016) Coordinated interaction between hippocampal sharp-wave ripples and anterior cingulate unit activity. J Neurosci 36:10663-10672
- Makino Y, Polygalov D, Bolaños F, Benucci A, McHugh TJ (2019) Physiological signature of memory age in the prefrontal-hippocampal circuit. Cell Rep 29:3835-3846.e5
- Goshen I, Brodsky M, Prakash R, Wallace J, Gradinaru V, Ramakrishnan C, Deisseroth K (2011) Dynamics of retrieval strategies for remote memories. Cell 147:678-689
- Frankland PW, Ding HK, Takahashi E, Suzuki A, Kida S, Silva AJ (2006) Stability of recent and remote contextual fear memory. Learn Mem 13:451-457
- Sierra-Mercado D, Padilla-Coreano N, Quirk GJ (2011) Dissociable roles of prelimbic and infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression and extinction of conditioned fear. Neuropsychopharmacology 36:529-538
- Corcoran KA, Quirk GJ (2007) Activity in prelimbic cortex is necessary for the expression of learned, but not innate, fears. J Neurosci 27:840-844
- Burgos-Robles A, Vidal-Gonzalez I, Quirk GJ (2009) Sustained conditioned responses in prelimbic prefrontal neurons are correlated with fear expression and extinction failure. J Neurosci 29:8474-8482
- Do-Monte FH, Quiñones-Laracuente K, Quirk GJ (2015) A temporal shift in the circuits mediating retrieval of fear memory. Nature 519:460-463
- Quiñones-Laracuente K, Vega-Medina A, Quirk GJ (2021) Time-dependent recruitment of prelimbic prefrontal circuits for retrieval of fear memory. Front Behav Neurosci 15:665116
- Park S, Kramer EE, Mercaldo V, Rashid AJ, Insel N, Frankland PW, Josselyn SA (2016) Neuronal allocation to a hippocampal engram. Neuropsychopharmacology 41:2987-2993
- Lau JMH, Rashid AJ, Jacob AD, Frankland PW, Schacter DL, Josselyn SA (2020) The role of neuronal excitability, allocation to an engram and memory linking in the behavioral generation of a false memory in mice. Neurobiol Learn Mem 174:107284
- Floresco SB, Tse MT (2007) Dopaminergic regulation of inhibitory and excitatory transmission in the basolateral amygdala-prefrontal cortical pathway. J Neurosci 27:2045-2057
- Kitamura T, Ogawa SK, Roy DS, Okuyama T, Morrissey MD, Smith LM, Redondo RL, Tonegawa S (2017) Engrams and circuits crucial for systems consolidation of a memory. Science 356:73-78
- Wheeler AL, Teixeira CM, Wang AH, Xiong X, Kovacevic N, Lerch JP, McIntosh AR, Parkinson J, Frankland PW (2013) Identification of a functional connectome for long-term fear memory in mice. PLoS Comput Biol 9:e1002853
- Silva BA, Burns AM, Gräff J (2019) A cFos activation map of remote fear memory attenuation. Psychopharmacology (Berl) 236:369-381
- Laurent V, Westbrook RF (2009) Inactivation of the infralimbic but not the prelimbic cortex impairs consolidation and retrieval of fear extinction. Learn Mem 16:520-529
- Awad W, Ferreira G, Maroun M (2015) Dissociation of the role of infralimbic cortex in learning and consolidation of extinction of recent and remote aversion memory. Neuropsychopharmacology 40:2566-2575
- Bayer H, Bertoglio LJ (2020) Infralimbic cortex controls fear memory generalization and susceptibility to extinction during consolidation. Sci Rep 10:15827
- Vidal-Gonzalez I, Vidal-Gonzalez B, Rauch SL, Quirk GJ (2006) Microstimulation reveals opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn Mem 13:728-733
- Bloodgood DW, Sugam JA, Holmes A, Kash TL (2018) Fear extinction requires infralimbic cortex projections to the basolateral amygdala. Transl Psychiatry 8:60
- Thompson BM, Baratta MV, Biedenkapp JC, Rudy JW, Watkins LR, Maier SF (2010) Activation of the infralimbic cortex in a fear context enhances extinction learning. Learn Mem 17:591-599
- Soler-Cedeño O, Cruz E, Criado-Marrero M, Porter JT (2016) Contextual fear conditioning depresses infralimbic excitability. Neurobiol Learn Mem 130:77-82
- Herry C, Garcia R (2002) Prefrontal cortex long-term potentiation, but not long-term depression, is associated with the maintenance of extinction of learned fear in mice. J Neurosci 22:577-583
- Milad MR, Quirk GJ (2002) Neurons in medial prefrontal cortex signal memory for fear extinction. Nature 420:70-74
- Marek R, Xu L, Sullivan RKP, Sah P (2018) Excitatory connections between the prelimbic and infralimbic medial prefrontal cortex show a role for the prelimbic cortex in fear extinction. Nat Neurosci 21:654-658
- Watanabe M, Uematsu A, Johansen JP (2021) Enhanced synchronization between prelimbic and infralimbic cortices during fear extinction learning. Mol Brain 14:175
- Mukherjee A, Caroni P (2018) Infralimbic cortex is required for learning alternatives to prelimbic promoted associations through reciprocal connectivity. Nat Commun 9:2727
- Silva BA, Astori S, Burns AM, Heiser H, van den Heuvel L, Santoni G, Martinez-Reza MF, Sandi C, Gräff J (2021) A thalamo-amygdalar circuit underlying the extinction of remote fear memories. Nat Neurosci 24:964-974
- Tao Y, Cai CY, Xian JY, Kou XL, Lin YH, Qin C, Wu HY, Chang L, Luo CX, Zhu DY (2021) Projections from infralimbic cortex to paraventricular thalamus mediate fear extinction retrieval. Neurosci Bull 37:229-241
- Kirouac GJ (2015) Placing the paraventricular nucleus of the thalamus within the brain circuits that control behavior. Neurosci Biobehav Rev 56:315-329
- Flavell SW, Greenberg ME (2008) Signaling mechanisms linking neuronal activity to gene expression and plasticity of the nervous system. Annu Rev Neurosci 31:563-590
- Xia Z, Dudek H, Miranti CK, Greenberg ME (1996) Calcium influx via the NMDA receptor induces immediate early gene transcription by a MAP kinase/ERK-dependent mechanism. J Neurosci 16:5425-5436
- Bast T, Zhang WN, Feldon J (2003) Dorsal hippocampus and classical fear conditioning to tone and context in rats: effects of local NMDA-receptor blockade and stimulation. Hippocampus 13:657-675
- Zhao MG, Toyoda H, Lee YS, Wu LJ, Ko SW, Zhang XH, Jia Y, Shum F, Xu H, Li BM, Kaang BK, Zhuo M (2005) Roles of NMDA NR2B subtype receptor in prefrontal long-term potentiation and contextual fear memory. Neuron 47:859-872
- Sacktor TC, Osten P, Valsamis H, Jiang X, Naik MU, Sublette E (1993) Persistent activation of the zeta isoform of protein kinase C in the maintenance of long-term potentiation. Proc Natl Acad Sci U S A 90:8342-8346
- Ko HG, Kim JI, Sim SE, Kim T, Yoo J, Choi SL, Baek SH, Yu WJ, Yoon JB, Sacktor TC, Kaang BK (2016) The role of nuclear PKMζ in memory maintenance. Neurobiol Learn Mem 135:50-56
- Li XY, Ko HG, Chen T, Descalzi G, Koga K, Wang H, Kim SS, Shang Y, Kwak C, Park SW, Shim J, Lee K, Collingridge GL, Kaang BK, Zhuo M (2010) Alleviating neuropathic pain hypersensitivity by inhibiting PKMzeta in the anterior cingulate cortex. Science 330:1400-1404
- Shum FW, Wu LJ, Zhao MG, Toyoda H, Xu H, Ren M, Pinaud R, Ko SW, Lee YS, Kaang BK, Zhuo M (2007) Alteration of cingulate long-term plasticity and behavioral sensitization to inflammation by environmental enrichment. Learn Mem 14:304-312
- Xue YX, Zhu ZZ, Han HB, Liu JF, Meng SQ, Chen C, Yang JL, Wu P, Lu L (2015) Overexpression of protein kinase Mζ in the prelimbic cortex enhances the formation of long-term fear memory. Neuropsychopharmacology 40:2487
- da Silva TR, Raymundi AM, Bertoglio LJ, Andreatini R, Stern CA (2020) Role of prelimbic cortex PKC and PKMζ in fear memory reconsolidation and persistence following reactivation. Sci Rep 10:4076
- Miao HH, Miao Z, Pan JG, Li XH, Zhuo M (2021) Brain-derived neurotrophic factor produced long-term synaptic enhancement in the anterior cingulate cortex of adult mice. Mol Brain 14:140
- Vetere G, Restivo L, Cole CJ, Ross PJ, Ammassari-Teule M, Josselyn SA, Frankland PW (2011) Spine growth in the anterior cingulate cortex is necessary for the consolidation of contextual fear memory. Proc Natl Acad Sci U S A 108:8456-8460
- Joseph NF, Zucca A, Wingfield JL, Espadas I, Page D, Puthanveettil SV (2021) Molecular motor KIF3B in the prelimbic cortex constrains the consolidation of contextual fear memory. Mol Brain 14:162
- Vetere G, Restivo L, Ammassari-Teule M (2012) Pre-synaptic control of remote fear extinction in the neocortex. Front Behav Neurosci 6:34
- Guenthner CJ, Miyamichi K, Yang HH, Heller HC, Luo L (2013) Permanent genetic access to transiently active neurons via TRAP: targeted recombination in active populations. Neuron 78:773-784
- Laroche S, Jay TM, Thierry AM (1990) Long-term potentiation in the prefrontal cortex following stimulation of the hippocampal CA1/subicular region. Neurosci Lett 114:184-190
- Taylor CJ, Ohline SM, Moss T, Ulrich K, Abraham WC (2016) The persistence of long-term potentiation in the projection from ventral hippocampus to medial prefrontal cortex in awake rats. Eur J Neurosci 43:811-822
- Park AJ, Harris AZ, Martyniuk KM, Chang CY, Abbas AI, Lowes DC, Kellendonk C, Gogos JA, Gordon JA (2021) Reset of hippocampal-prefrontal circuitry facilitates learning. Nature 591:615-619
- Vieira PA, Corches A, Lovelace JW, Westbrook KB, Mendoza M, Korzus E (2015) Prefrontal NMDA receptors expressed in excitatory neurons control fear discrimination and fear extinction. Neurobiol Learn Mem 119:52-62
- Burgos-Robles A, Vidal-Gonzalez I, Santini E, Quirk GJ (2007) Consolidation of fear extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex. Neuron 53:871-880
- Soler-Cedeño O, Torres-Rodríguez O, Bernard F, Maldonado L, Hernández A, Porter JT (2019) Plasticity of NMDA receptors at ventral hippocampal synapses in the infralimbic cortex regulates cued fear. eNeuro 6:ENEURO.0354-18.2019
- Garín-Aguilar ME, Díaz-Cintra S, Quirarte GL, Aguilar-Vázquez A, Medina AC, Prado-Alcalá RA (2012) Extinction procedure induces pruning of dendritic spines in CA1 hippocampal field depending on strength of training in rats. Front Behav Neurosci 6:12
- Choi DI, Kim J, Lee H, Kim JI, Sung Y, Choi JE, Venkat SJ, Park P, Jung H, Kaang BK (2021) Synaptic correlates of associative fear memory in the lateral amygdala. Neuron 109:2717-2726.e3
- Vetere G, Restivo L, Novembre G, Aceti M, Lumaca M, Ammassari-Teule M (2011) Extinction partially reverts structural changes associated with remote fear memory. Learn Mem 18:554-557
- Laricchiuta D, Sciamanna G, Gimenez J, Termine A, Fabrizio C, Caioli S, Balsamo F, Panuccio A, De Bardi M, Saba L, Passarello N, Cutuli D, Mattioni A, Zona C, Orlando V, Petrosini L (2021) Optogenetic stimulation of prelimbic pyramidal neurons maintains fear memories and modulates amygdala pyramidal neuron transcriptome. Int J Mol Sci 22:810
- Duvarci S, Nader K, LeDoux JE (2005) Activation of extracellular signal-regulated kinase- mitogen-activated protein kinase cascade in the amygdala is required for memory reconsolidation of auditory fear conditioning. Eur J Neurosci 21:283-289
- Fukushima H, Zhang Y, Kida S (2021) Active transition of fear memory phase from reconsolidation to extinction through ERK-mediated prevention of reconsolidation. J Neurosci 41:1288-1300
- Khalaf O, Gräff J (2019) Reactivation of recall-induced neurons in the infralimbic cortex and the basolateral amygdala after remote fear memory attenuation. Front Mol Neurosci 12:70