Articles

Article Tools
 View Full Text View Full Text |
 Abstract Abstract |
 Article as PDF Article as PDF |
 Print this Article Print this Article |
 Pubmed Pubmed |
 PMC PMC |
 PubReader PubReader |
 Export to Citation Export to Citation |
 Email Alerts Email Alerts |
 Open Access Open Access |
Share this article on :



Stats or Metrics
Article
Review Article
Exp Neurobiol 2016; 25(5): 241-251
Published online October 31, 2016
https://doi.org/10.5607/en.2016.25.5.241
© The Korean Society for Brain and Neural Sciences
Inflammation after Ischemic Stroke: The Role of Leukocytes and Glial Cells
Jong Youl Kim1, Joohyun Park1,2, Ji Young Chang1, Sa-Hyun Kim3 and Jong Eun Lee1,2*
1Department of Anatomy, Yonsei University College of Medicine, 2Bk21 Plus Project for Medical Sciences and Brain Research Institute, Yonsei University College of Medicine, Seoul 03722, 3Department of Clinical Laboratory Science, Semyung University, Jaecheon 27136, Korea
Correspondence to: *To whom correspondence should be addressed.
TEL: 82-2-2228-1646, 1659, FAX: 82-2-365-0700
e-mail: jelee@yuhs.ac
Abstract
The immune response after stroke is known to play a major role in ischemic brain pathobiology. The inflammatory signals released by immune mediators activated by brain injury sets off a complex series of biochemical and molecular events which have been increasingly recognized as a key contributor to neuronal cell death. The primary immune mediators involved are glial cells and infiltrating leukocytes, including neutrophils, monocytes and lymphocyte. After ischemic stroke, activation of glial cells and subsequent release of pro- and anti-inflammatory signals are important for modulating both neuronal cell damage and wound healing. Infiltrated leukocytes release inflammatory mediators into the site of the lesion, thereby exacerbating brain injury. This review describes how the roles of glial cells and circulating leukocytes are a double-edged sword for neuroinflammation by focusing on their detrimental and protective effects in ischemic stroke. Here, we will focus on underlying characterize of glial cells and leukocytes under inflammation after ischemic stroke.
Keywords: ischemic stroke, inflammation, glial cells, leukocytes
INTRODUCTION
The inflammatory response following acute ischemic stroke is a well-known and widely studied phenomenon. Pathological features of ischemia such as necrotic cells, cell death debris, and increased reactive oxygen species (ROS) can induce neuroinflammatory by activating resident microglia and astrocytes as well as attracting infiltrating leukocytes from circulating blood. The recruitment of both brain and peripheral immune cell types in post-ischemic tissue can accelerate and expand an infarct initiated by ischemic insult.
The first-line responders to central nervous system (CNS) injury are microglia and astrocytes. Microglia are the resident macrophages of the brain and a key modulator of immunologic responses after ischemic stroke [1,2]. Microglia constitute 15% of the total glial cell population in the adult murine brain (16.6% in humans) and are primarily found in brain gray matter [3]. Once activated by extracellular signals, they function to sweep debris and toxic substances by phagocytosis, thereby helping maintain normal cellular homeostasis in the brain [4,5]. Activated microglia also increase secretion of cytokines and leukocyte adhesion molecules within cerebral vasculature, all within 24 hours of the ischemic insult [6,7]. Astrocytes are the most abundant cells in the brain. In uninjured brain tissues, astrocytes provide structural and nutritive support for neurons. After ischemic stroke, astrocytes play an important role in wound healing and repair by mediating reactive gliosis and glial scar formation [8]. Both astrocytes and microglia may also produce inflammatory cytokines and toxic mediators such as excitotoxic glutamate.
Within a couple days to a week after ischemic stroke, peripheral circulating leukocytes are also recruited to the injury response. Neutrophils, monocytes, and lymphocytes infiltrate the CNS by binding to adhesion molecules on activated endothelial cells. Activated endothelia further enhance adhesion binding of circulating leukocytes by causing microvascular occlusion and infiltration of immune cells into the brain parenchyma [9,10]. Once in the CNS, leukocytes release inflammatory cytokines at the site of injury such as tumor necrosis factor-α (TNF-α), interleukin-1 beta (IL-1β) and interleukin-6 (IL-6) [11]. Several studies have shown that mechanism of leukocyte infiltration could be a possible way to widen the therapeutic window after ischemic stroke [12,13,14]. Given that the immune system consists of a variety of different cell populations with a range of different functions at different time points following tissue damage, detailed analyses of specific immune cell infiltration is certainly needed.
This review examines the role of glia and leukocytes in ischemic stroke, exploring how these two sets of immune cells work in tandem to both mediate repair and augment injury. Better understanding these mechanisms may help expand stroke's narrow therapeutic window and lead to the discovery of novel pharmacologic interventions involving one or more immune cell population(s).
DYNAMIC MICROGLIA AND ASTROCYTE IN ISCHEMIC STROKE
Microglia are key modulators of the immune response in the brain and are considered the resident immune cell of the central nervous system. Under normal conditions, microglia are primarily involved in activity-dependent synaptic pruning and repair [15]. In the event of acute brain injury, microglia can quickly undergo morphologic transformation from a ramified resting state, characterized by many branching processes, to an active, motile ameboid state, where they become virtually indistinguishable from circulating macrophages [16,17]. Therefore, activated microglia are often called brain macrophages. Active microglia can then phagocytose foreign organisms as well as injured brain cells [18,19,20]. In ischemic stroke, microglial activation occurs in the early stages of neuroinflammation; activated microglia can be detected in lesions as early as 2 hours post-ischemia and can be detected up to 1 week after brain injury [21]. Several reports have demonstrated that the direct application of activated microglia has been shown to effect cell death in neurons [22,23]. In
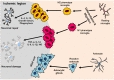
In experimental stroke models, activated microglia have been shown to migrate toward the ischemic hemisphere of the cerebral cortex [29]. The precise mechanisms of microglial activation following ischemia are unclear. However, the literature to date strongly indicates that activate microglia have predominantly harmful effects in the acute stages of ischemic stroke and that most beneficial effects appear in delayed stages. Numerous studies indicate that different signals lead to two primary activation phenotypes: classically activated (M1) and alternatively activated (M2) [30,31,32]. The M1 phenotype microglia, activated by lipopolysaccharide (LPS) and the proinflammatory cytokine interferonc (IFNc), induces transcriptional activation of nuclear factor-kB and makes high levels of proinflammatory cytokines and oxidative metabolites such as TNF-α, interleukin (IL)-12, IL-6, IL-1β, and nitric oxide (NO), formerly indicated to cause additional damage (Fig. 1) [33]. In contrast, the M2 phenotype microglia is promoted by anti-inflammatory cytokines such as IL-4 or IL-13 [34], which are considered to prevent inflammation and improve tissue repair and wound healing (Fig. 1) [35]. Microglial activation may begin with an M1 phenotype which mediates an innate or an adaptive immune response and ultimately exacerbates neuronal damage [36,37]. At later timepoints after injury, microglia may also transform the M2 phenotype microglia which facilitate repairoriented functions by secreting growth factors such as vascular endothelial growth factor or brain-derived neurotrophic factor, and by clearing cellular debris via phagocytosis. M2 phenotype microglia tend to limit proinflammatory signal production. The timing and mechanism of M1 vs M2 phenotype activation is an important element to consider when manipulating microglia in stroke, and it is important to identify and understand the different microglial phenotypes and their unique functions at different time points [38,39].
Recent research has also demonstrated that microglia can switch from the M1 phenotype to the M2 phenotype [40,41,42]. One such study showed that HIV-associated dementia initiates and maintains M1 phenotype microglia in the event of CD40 ligation by CD40L and TNF. These glia may later switch glia to the M2 phenotype via upregulation of CD45 [43]. In another study of aged mice subjected to brain injury, histological results indicated that aged brains of injured mice had not only larger lesions and worsened outcome, but also showed microglial polarization toward M1 phenotype compared to younger mice [44].
The dual phenotype functions make microglia a promising target for treating ischemic stroke. Future research for stroke therapies can build on studies characterizing different modes of microglia activation by, for example, dampening injury-exacerbating M1 phenotype functions or by inducing or amplifying native M2 phenotype repair functions at the appropriate time intervals following injury. For example, one recent study showed that activating microglia membrane protein triggering receptor expressed on myeloid cells 2 (TREM2) may also lead to phagocytic activity [45] and drive other anti-inflammatory functions [46]. Transgenic TREM2-deficient mice showed poorer recovery following ischemic stroke [47]. The challenge will be to discover methods for selectively suppressing the detrimental effects of microglial activation without compromising the restorative properties such as repair and remodeling. Studies in therapeutic microglial targets will also need to find ways to suppress cytotoxic mechanisms without disrupting beneficial effects. In neurodegenerative diseases, down-regulating CD40 or turning on CD45 has been shown to induce M2 phenotype and improve neurological outcome [43]. Similar strategies may also prove to protect against brain ischemia.
Astrocytes are important mediators of homeostasis in the brain, including the regulation of immune reactions. In addition to their immunological functions, astrocytes have been reported to release various pro-inflammatory factors after ischemic injury, such as glial fibriliary acidid protein (GFAP) [48]. These cells also play an central role in enhancing reactive gliosis and glial scar formation (Fig. 1) [8]. Although an important part of the long-term healing, astrocytic gliosis may also be destructive after brain injury [18]. A massive astroglial response appears in the core of the lesion from hour 4 to day 1 following ischemic stroke, and reaches a peak around day 4 [49]. This glial scar has both neurotoxic and neurotrophic properties. The scar acts as a barrier which prevents axonal ingrowth and reinnervation, thus interrupting recovery. However, this scar also isolates damaged tissue from viable tissue and prevents additional damage to the surrounding brain [49]. Recent study showed beneficial effect of astrocytic gliosis which aids rather than prevents CNS axon regeneration [50]
In addition to modulating scar formation, astrocytes have also been observed releasing various immune molecules such as cytokines, chemokines and inducible nitric oxide synthase (iNOS), and inducing a Th2 (anti-inflammatory) immune response [51]. In ischemic stroke, iNOS was observed in reactive hippocampal astrocytes [52]. The inflammatory role of astrocytes has also been demonstrated in a study of TNF-like weak inducer of apoptosis (TWEAK), a member of the TNF superfamily. TWEAK was detected on neurons, astrocytes and endothelial cells, where it was shown to increase pro-inflammatory molecule production through interaction with the astrocytic Fn14 receptor [53,54]. TWEAK and Fn14 expression have been documented in experimental stroke models, where Fn14 inhibition led to decreased ischemic injury [54]. These results show that while astrocytes have long been viewed to play scaffolding and supportive roles for neurons, activated astrocytes may also be detrimental in the ischemic brain, not unlike microglia and other immune cells.
INFILTRATED LEUKOCYTES EXACERBATE ISCHEMIC STROKE
Infiltrated leukocytes promote cerebral ischemic injury in a number of different ways. First, adhesion of leukocytes to the endothelium can reduce the flow of erythrocytes through the microvasculature causing the cerebral no-reflow phenomenon and additional brain injury. Activated leukocytes at the surface of the endothelium also release proteases, ROS, gelatinases, and collagenases, and impair potentially salvageable blood vessels and brain tissues. Phospholipase activation in leukocytes leads to a production of biologically active substances, such as leukotrienes, eicosanoids, prostaglandins, and platelet-activating factor, which result in vasoconstriction and extend platelet aggregation. Lastly, infiltrated leukocytes release pro-inflammatory cytokines and other immune modulators in the penumbra surrounding the infarct core causing further neuronal injury [55,56,57,58].
Neutrophils are the first blood-borne immune cells to arrive at ischemic brain tissues. These innate immune cells have important roles in acute ischemic brain injury and in the events leading up to infarction such as atherosclerosis and thrombus formation. Following ischemic stroke, neutrophils may cause sterile inflammation by interacting with endothelial adhesion molecules to slow their intravascular movement and induce polarization, which causes adhesion to the pro-inflammatory endothelium [59]. Neutrophils attach to the endothelium by binding various adhesion molecules including the selectins (P-, E-, and L-selectin), intracellular cell adhesion molecule-1 (ICAM-1) and integrins (CD11a, b and c) within 15 minutes of ischemic stroke [60,61]. By 2 hours, neutrophils rolling and adhesion appear in the pial vessels of brain [62]. After initial adherence, neutrophils will follow a chemokine and activator gradient produced by the injured tissue. Neutrophils reach peak numbers at 2~4 days after ischemic stroke and then decrease thereafter [63,64]. During this period, pro-inflammatory neutrophil activation contributes to disruption of blood brain barrier, increased infarct size, hemorrhagic transformation, and worse neurologic outcomes.
Neutrophil adhesion is an important step in the immune response to ischemic brain injury [65]. Adhesion molecules attach immune cells tightly to the endothelial wall, thereby stimulating and facilitating diapedesis through the vessel wall to the site of ischemic brain injury [65]. These neutrophil adhesion factors include ICAM-1, MAC-1 (CD11b/CD18), and selectins. Expression of ICAM-1 is increased in endothelia proximal to the injured brain within hours after stroke onset, and peaks at about 12-48 hours [66]. Mice deficient in adhesion molecules either by transgenic manipulation or pharmacologic interruption of ICAM-1 activity have been shown with decreased infarct areas and reduced brain leukocyte infiltration in experimental stroke [67,68]. Neutrophils express CD11b/CD18, also known as MAC-1, which are integrins that contain a common β2 chain (CD18) and are thus classified as β2 integrins. Integrins are located in neutrophil plasma membranes, where they bind to endothelial ICAM-1 and enable cell migration through the vessel wall. In MAC-1 deficient transgenic mice, infarct size, mortality, and neutrophil infiltration into ischemic brain are reduced after ischemic stroke [69]. Another study inhibited MAC-1 via monoclonal antibodies in experimental stroke models and found decreased infarct sizes and improved functional outcomes in rabbits [70]. Similarly, blocking MAC-1 with recombinant neutrophil inhibitory factor (rNIF) is associated with reduced infarct size, and improved neurological outcomes [71]. However, this benefit only occurs when rNIF is administered early after cerebral reperfusion (2 to 6 hours) and is not effective in models where no reperfusion occurs. Lastly, selectins are calcium-dependent, transmembrane glycoproteins that bind to carbohydrate residues (sialyl-Lewis X), and mediate rolling and adhesion to vascular endothelium. E-, P-, and L-selectin work together to coordinate neutrophil trafficking after brain ischemia [72]: E-selectin and P-selectin participate in initial neutrophils rolling and recruitment [73], whereas L-selectin guides unstimulated neutrophils to areas of activated endothelium [74]. Various experimental stroke models have found positive correlations between P- and E-selectin upregulation and the promotion of post-ischemic inflammatory responses as well as injury severity [75,76].
Inflammatory neutrophil activity after ischemic stroke is further modulated by interactions with chemokines and other cytokines, both of which are released in brain tissues after injury. These signaling ligands can increase the neutrophil recruitment and migration mechanisms mediated by adhesion factors by binding and activating neutrophils. Chemokine receptors are expressed on neutrophil plasma membranes, and their activation drives neutrophils to sustain or even amplify inflammatory pathways at brain lesion. Different classes of chemokines are identified by their structures, the main classes being CXC, CC, XC and CX3C. The "C"s refer to N-terminal cysteine residues, and the classes are divided depending on whether these residues flank an amino acid between them (CXC) or whether they are adjacent (CC). The different classes of chemokines act through unique and overlapping receptors which are a part of a superfamily of G-protein-coupled receptors [77]. The CXC class can be further split into ELR+ or ELR- groups based on whether the glutamate-leucine-arginine motif is present between the N-terminus and the first cysteine [78]. Chemokine receptors can also be flexible, binding multiple classes or subtypes of ligands, and many different chemokines are capable of activating neutrophils [79]. However, the ELR+ CXC chemokine subfamily are thought to be mainly neutrophil chemoattractants, whereas the CC chemokines more typically attract monocytes and T lymphocytes [80]. Though as a whole, their signaling is associated with more general pro-inflammatory mechanisms and pathways, several chemokines in CXC group have also been directly implicated in mediating neutrophil infiltration [81]. Because of this, chemokine signaling is closely associated with worse stroke outcomes [60], and chemokine ligands and receptors have become a hot topic of research investigating potential therapeutic targets. One recent study showed large increases in expression of key members of the ELR+ CXC chemokine subfamily, the neutrophil receptor CXCR2, and its ligands CXCL1 and CXCL2, which reached maximum levels at 1 to 3 days after injury [81]. In the rodent stroke model, inhibition of CXCR1 and CXCR2 with Reparixin decreased infarct size, improved motor function, and reduced brain levels of MPO and IL-1β [82].
Monocytes are derived from hematopoietic stem cells (HSC) in the liver and spleen during embryonic development and primarily in the bone marrow after birth [83,84]. Like neutrophils, monocytes are incompletely differentiated cells that have a highly phagocytic capacity and react depending on the nature of stimuli within their microenvironment [84,85]. By expression of specific surface markers, this cell type can be divided into pro-inflammatory (classically activated) or anti-inflammatory (alternatively activated) subsets. Recruitment of circulating monocytes to the ischemic brain is similar to that of neutrophils, orchestrated by inflammatory cytokines, such as adhesion molecules and chemokines. In stroke models, the monocyte chemoattractant protein-1 (MCP-1, CCL2) and its receptor CCR2 are known to be involved in the inflammatory response of the injured brain [86]. At baseline, CCL2 mRNA expression is almost absent, but ischemia leads to a significant increase in MCP-1 mRNA expression in injured portions of the cortex after either permanent or temporary MCA occlusion around 12 h to 2 days and remained elevated up to 5 days [87,88]. Pro-inflammatory monocytes express CCR2 with low or no expression of CX3CR1 across different species. CCR2 expression is critical for the trafficking of circulating monocytes into injured brain where they can transform into macrophages. Anti-inflammatory monocytes, which do not express CCR2 but do express higher levels of CX3CR1, patrol blood vessels in a steady state and perform in situ phagocytosis [85,89]. Several other studies showed that CCL2 or CCR2 deficient mice reduce phagocytic macrophage accumulation with smaller infarcts in experimental stroke models, suggesting CCR2 monocytes may have a deleterious effect [90,91]. In rodents, monocytes fall into two main subsets based on chemokine receptor and Ly-6C (Gr-1) expression levels. Ly6Chigh pro-inflammation has a short half-life and is actively recruited to inflamed tissues, contributing to the inflammatory response. Ly6Clow anti-inflammatory has a longer half-life and is the subtype responsible for patrolling the lumen of blood vessels, contributing to the maintenance of vascular homeostasis [92]. One recent paper demonstrated an increase in Ly6Chigh monocytes at day 3 after stroke, whereas the number of Ly6Clow monocytes was greatest at day 6, paralleled by sequential peaks of CCR2 and CX3CR1 mRNA as well as gene expression of the pro- and anti-inflammatory cytokines IL-1β and TGF-β, respectively [93]. These data may indicate the presence of a dynamic shift in the recruitment and infiltration of monocyte subsets into injured brain after ischemic stroke, or probably the differentiation of pro-inflammatory monocytes (Ly6Chigh) into anti-inflammatory monocytes (Ly6Clow). Better understanding the mechanisms mediating differentiation into these two monocyte subsets may reveal additional therapeutic strategies for controlling inflammation after ischemic injury.
The patterns and consequences of lymphocyte activity after stroke are not as well characterized as that of neutrophils and monocytes. A number of studies have observed that lymphocytes may negatively contribute to brain injury pathogenesis. Similar to neutrophils and monocytes, these cells also release pro-inflammatory cytokines and cytotoxic substances, such as ROS. Several stroke studies have shown that the number of lymphocytes spikes in the ischemic brain at later time-points than neutrophils [94,95]. Interrupting lymphocyte entry into infarcted brain tissues decreases the severity of the injury, and suggests that, like neutrophils, lymphocytes perform an overall harmful role [96]. T lymphocytes are a key player in amplifying inflammation after ischemic stroke, whereas B lymphocytes have been shown to play a lesser role [81]. One study showed that T lymphocyte-deficient mice had smaller infarct sizes and improved neurological outcomes relative to control groups in a model of transient focal ischemia [13,97]. This and other studies of lymphocyte-deficient mice together suggest that the neuroprotective effects produced by lymphocyte suppression in stroke appear dependent on the absence of T lymphocytes and not B lymphocytes because the reconstitution of B lymphocytes does not affect the protection observed. By contrast, when T lymphocytes are transplanted back in to Rag1-deficient mice, this protection disappeared [13,97,98]. This distinction between T and B lymphocytes remains somewhat controversial since another recent study failed to see noteworthy differences in infarct size between immune-deficient mice (deficient in both T and B lymphocytes) and wildtype after stroke [99]. The cause for these conflicting observations is unclear, although they may be related to the type of stroke; the latter study adopted a permanent model of focal cerebral ischemia instead of the temporary focal ischemia model used in the first study.
Another faction of the current literature suggests that not all T lymphocytes subtypes are detrimental to acute brain injured outcome. In one study that recreated ischemia-like injury in cultured primary neurons, isolated neutrophils released excitotoxins that induced neuronal death, whereas lymphocytes were not shown to be neurotoxic and actually increased astrocyte proliferation [100]. Another group showed that natural killer (NK) cells and T lymphocyte functions that rely on T cell receptor co-activation may not influence ischemic injury at all [98]. Furthermore, the impact of regulatory T (Treg) lymphocytes is still in question. Liesz et al. reported that infarct volume and neurological deficit were significantly increased in mice given an antibody to neutralize Treg lymphocytes compared to controls Beneficial effects of Treg in brain ischemia were mediated by IL-10 [101]. These data also demonstrated that IL-10 may be important for this immunomodulatory event. However, other study could not show any modulatory role of Treg cells [102]. While there is little evidence of their impact at the site of ischemic injury, NK cells may nevertheless influence stroke patient outcomes. In the liver, resident NK cell function is reduced by augmented sympathetic neurotransmission, which leads to the immunosuppression and susceptibility to infections that occur following stroke [103].
Whether harmful or protective, the mechanisms of lymphocyte activity after stroke are currently unclear. However, among the various theorized roles of lymphocytes in stroke, injury-exacerbating mechanisms of T lymphocytes have been best characterized to date. Generally, T lymphocytes attack bacteriaand virus-infected cells by releasing cytokines or cytotoxins [104], and they may respond similarly at sterile ischemic brain lesions. These T lymphocytes secretions cause cell death via interaction with the Fas receptor [105], and a few studies reported that neutralization of T lymphocyte-derived cytokines (IL-17, IL-12, IL-23, interferon gamma) reduced brain injury and improved neurological outcomes in experimental stroke models [13,101]. Another paper showed that mice lacking T lymphocyte-secreted perforin demonstrated significant neuroprotection, suggesting another pathway by which lymphocytes may contribute to ischemic damage [48]. In addition to these findings, yet another recent study observed circulating T cells producing 7 to 15 fold greater amounts of NADPH oxidase type 2 (Nox2)-derived superoxide after ischemic stroke, suggesting that peripheral T lymphocytes may exacerbate oxidative tissue injury at the brain lesion remotely, without having to migrate to the brain lesion, by releasing Nox-2-derived superoxide into the blood [106].
CONCLUSIONS
The role of inflammation following ischemic stroke has become an increasingly popular area for understanding interactions between the peripheral immune system and brain injury. Glia and leukocytes are considered the two major classes of immunocompetent cells involved in ischemic brain injury, and their activation and recruitment represent key stages in in initiating and sustaining neuroinflammation. However, our understanding of the mechanisms governing their activation and function after ischemia is still limited. For this reason, much effort has been directed toward an understanding of where each cell migrates and localizes in the ischemic brain, when they accumulate and infiltrate in brain injury, and how they become activated and mediate neuroinflammation (Table 1). Future studies investigating the spatial, temporal, and functional attributes of glia and leukocytes are necessary for greater insight into how these cells may exacerbate or protect against ischemic stroke. The mechanisms underlying these functions are critical to understand for developing new therapeutic strategies to treat acute ischemic brain.
Figures
{kind=link}
Tables
{kind=link}
| Cell type | Major function(s) | Reference(s) |
|---|---|---|
| Microgliae | - activity-dependent synaptic pruing and repair | [15] |
| - morphologic transformation in acute brain injury | [16,17] | |
| - sweep foregin organism in brain injury | [18,19,20], [21] | |
| - role of M1 phenotype microglia in the brain | [25], [28,29] | |
| - role of M2 phenotype microglia in the brain | [26], [27], [30,31] | |
| - swith of M1 to M2 microglia | [32,33,34], [35], [36] | |
| - microglia functionin ischemic stroke | [37], [38,39], [40,41,42], [45], [46], [47] | |
| Astrocyte | - regulation of immune reactions | [48] |
| - reactive gliosis and glial scar formation | [8], [18], [49] | |
| - releasing various immune molecules | [50], [51], [52,53] | |
| Neutrophil | - roles in acute ischemic brain injury | [58] |
| - interaction with adhesive molecules | [59,60] | |
| - rolling and adhesion the pial vessels of brain | [61] | |
| - infiltration and immune response to ischemic brain | [70], [71], [72,73] | |
| Monocyte | - phagocytic capacity | [83,84] |
| - expression of chemokines | [85], [86,87] | |
| - expression of the pro- and anti-inflammatory cytokines | [92] | |
| T lymphocyte | - improvement of inflammation in ischemic brain | [68], [93,94], [95] |
| - increased astrocyte proliferation | [102] | |
| - oxidative tissue injury | [105] |
References
- Kreutzberg GW. Microglia: a sensor for pathological events in the CNS. Trends Neurosci 1996;19:312-318.

- Kim JY, Kawabori M, Yenari MA. Innate inflammatory responses in stroke: mechanisms and potential therapeutic targets. Curr Med Chem 2014;21:2076-2097.
- Aisen PS, Schmeidler J, Pasinetti GM. Randomized pilot study of nimesulide treatment in Alzheimer's disease. Neurology 2002;58:1050-1054.
- Mittelbronn M, Dietz K, Schluesener HJ, Meyermann R. Local distribution of microglia in the normal adult human central nervous system differs by up to one order of magnitude. Acta Neuropathol 2001;101:249-255.
- Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci 2007;10:1387-1394.
- Becker KJ. Inflammation and acute stroke. Curr Opin Neurol 1998;11:45-49.
- Stanimirovic DB, Wong J, Shapiro A, Durkin JP. Increase in surface expression of ICAM-1, VCAM-1 and E-selectin in human cerebromicrovascular endothelial cells subjected to ischemia-like insults. Acta Neurochir Suppl 1997;70:12-16.
- Pekny M, Nilsson M. Astrocyte activation and reactive gliosis. Glia 2005;50:427-434.
- Suzuki H, Abe K, Tojo S, Morooka S, Kimura K, Mizugaki M, Itoyama Y. Postischemic expression of P-selectin immunoreactivity in rat brain. Neurosci Lett 1997;228:151-154.
- Hallenbeck JM. Significance of the inflammatory response in brain ischemia. Acta Neurochir Suppl 1996;66:27-31.
- Tuttolomondo A, Di Raimondo D, di Sciacca R, Pinto A, Licata G. Inflammatory cytokines in acute ischemic stroke. Curr Pharm Des 2008;14:3574-3589.
- Iadecola C, Alexander M. Cerebral ischemia and inflammation. Curr Opin Neurol 2001;14:89-94.
- Yilmaz G, Arumugam TV, Stokes KY, Granger DN. Role of T lymphocytes and interferon-gamma in ischemic stroke. Circulation 2006;113:2105-2112.
- Hallenbeck JM, Hansson GK, Becker KJ. Immunology of ischemic vascular disease: plaque to attack. Trends Immunol 2005;26:550-556.
- El Khoury J, Hickman SE, Thomas CA, Loike JD, Silverstein SC. Microglia, scavenger receptors, and the pathogenesis of Alzheimer's disease. Neurobiol Aging 1998;19:S81-S84.
- Saijo K, Glass CK. Microglial cell origin and phenotypes in health and disease. Nat Rev Immunol 2011;11:775-787.
- Appel SH, Zhao W, Beers DR, Henkel JS. The microglial-motoneuron dialogue in ALS. Acta Myol 2011;30:4-8.
- Wang Q, Tang XN, Yenari MA. The inflammatory response in stroke. J Neuroimmunol 2007;184:53-68.
- Lai AY, Todd KG. Microglia in cerebral ischemia: molecular actions and interactions. Can J Physiol Pharmacol 2006;84:49-59.
- Schubert P, Morino T, Miyazaki H, Ogata T, Nakamura Y, Marchini C, Ferroni S. Cascading glia reactions: a common pathomechanism and its differentiated control by cyclic nucleotide signaling. Ann N Y Acad Sci 2000;903:24-33.
- Kawabori M, Yenari MA. The role of the microglia in acute CNS injury. Metab Brain Dis 2015;30:381-392.
- Huang WC, Qiao Y, Xu L, Kacimi R, Sun X, Giffard RG, Yenari MA. Direct protection of cultured neurons from ischemia-like injury by minocycline. Anat Cell Biol 2010;43:325-331.
- Lehnardt S, Massillon L, Follett P, Jensen FE, Ratan R, Rosenberg PA, Volpe JJ, Vartanian T. Activation of innate immunity in the CNS triggers neurodegeneration through a Toll-like receptor 4-dependent pathway. Proc Natl Acad Sci U S A 2003;100:8514-8519.
- Lee SC, Liu W, Roth P, Dickson DW, Berman JW, Brosnan CF. Macrophage colony-stimulating factor in human fetal astrocytes and microglia. Differential regulation by cytokines and lipopolysaccharide, and modulation of class II MHC on microglia. J Immunol 1993;150:594-604.
- Sawada M, Kondo N, Suzumura A, Marunouchi T. Production of tumor necrosis factor-alpha by microglia and astrocytes in culture. Brain Res 1989;491:394-397.
- Ritzel RM, Patel AR, Grenier JM, Crapser J, Verma R, Jellison ER, McCullough LD. Functional differences between microglia and monocytes after ischemic stroke. J Neuroinflammation 2015;12:106.
- Kriz J. Inflammation in ischemic brain injury: timing is important. Crit Rev Neurobiol 2006;18:145-157.
- Zhao BQ, Wang S, Kim HY, Storrie H, Rosen BR, Mooney DJ, Wang X, Lo EH. Role of matrix metalloproteinases in delayed cortical responses after stroke. Nat Med 2006;12:441-445.
- Yu YM, Kim JB, Lee KW, Kim SY, Han PL, Lee JK. Inhibition of the cerebral ischemic injury by ethyl pyruvate with a wide therapeutic window. Stroke 2005;36:2238-2243.
- Chawla A. Control of macrophage activation and function by PPARs. Circ Res 2010;106:1559-1569.
- Geissmann F, Gordon S, Hume DA, Mowat AM, Randolph GJ. Unravelling mononuclear phagocyte heterogeneity. Nat Rev Immunol 2010;10:453-460.
- Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol 2005;5:953-964.
- Nguyen KD, Qiu Y, Cui X, Goh YP, Mwangi J, David T, Mukundan L, Brombacher F, Locksley RM, Chawla A. Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis. Nature 2011;480:104-108.
- Nair MG, Guild KJ, Artis D. Novel effector molecules in type 2 inflammation: lessons drawn from helminth infection and allergy. J Immunol 2006;177:1393-1399.
- Colton CA. Heterogeneity of microglial activation in the innate immune response in the brain. J Neuroimmune Pharmacol 2009;4:399-418.
- Wang G, Zhang J, Hu X, Zhang L, Mao L, Jiang X, Liou AK, Leak RK, Gao Y, Chen J. Microglia/macrophage polarization dynamics in white matter after traumatic brain injury. J Cereb Blood Flow Metab 2013;33:1864-1874.
- Hu X, Li P, Guo Y, Wang H, Leak RK, Chen S, Gao Y, Chen J. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke 2012;43:3063-3070.
- Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol 2008;8:958-969.
- Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 2004;25:677-686.
- Morganti JM, Riparip LK, Rosi S. Call off the dog(ma): M1/M2 polarization is concurrent following traumatic brain injury. PLoS One 2016;11:e0148001.
- Yu Z, Sun D, Feng J, Tan W, Fang X, Zhao M, Zhao X, Pu Y, Huang A, Xiang Z, Cao L, He C. MSX3 switches microglia polarization and protects from inflammationinduced demyelination. J Neurosci 2015;35:6350-6365.
- Peferoen LA, Vogel DY, Ummenthum K, Breur M, Heijnen PD, Gerritsen WH, Peferoen-Baert RM, van der Valk P, Dijkstra CD, Amor S. Activation status of human microglia is dependent on lesion formation stage and remyelination in multiple sclerosis. J Neuropathol Exp Neurol 2015;74:48-63.
- Salemi J, Obregon DF, Cobb A, Reed S, Sadic E, Jin J, Fernandez F, Tan J, Giunta B. Flipping the switches: CD40 and CD45 modulation of microglial activation states in HIV associated dementia (HAD). Mol Neurodegener 2011;6:3.
- Kumar A, Stoica BA, Sabirzhanov B, Burns MP, Faden AI, Loane DJ. Traumatic brain injury in aged animals increases lesion size and chronically alters microglial/macrophage classical and alternative activation states. Neurobiol Aging 2013;34:1397-1411.
- Hsieh CL, Koike M, Spusta SC, Niemi EC, Yenari M, Nakamura MC, Seaman WE. A role for TREM2 ligands in the phagocytosis of apoptotic neuronal cells by microglia. J Neurochem 2009;109:1144-1156.
- Piccio L, Buonsanti C, Mariani M, Cella M, Gilfillan S, Cross AH, Colonna M, Panina-Bordignon P. Blockade of TREM-2 exacerbates experimental autoimmune encephalomyelitis. Eur J Immunol 2007;37:1290-1301.
- Kawabori M, Kacimi R, Kauppinen T, Calosing C, Kim JY, Hsieh CL, Nakamura MC, Yenari MA. Triggering receptor expressed on myeloid cells 2 (TREM2) deficiency attenuates phagocytic activities of microglia and exacerbates ischemic damage in experimental stroke. J Neurosci 2015;35:3384-3396.
- Benveniste EN. Cytokine actions in the central nervous system. Cytokine Growth Factor Rev 1998;9:259-275.
- Nowicka D, Rogozinska K, Aleksy M, Witte OW, Skangiel-Kramska J. Spatiotemporal dynamics of astroglial and microglial responses after photothrombotic stroke in the rat brain. Acta Neurobiol Exp(Warsz) 2008;68:155-168.
- Anderson MA, Burda JE, Ren Y, Ao Y, O'Shea TM, Kawaguchi R, Coppola G, Khakh BS, Deming TJ, Sofroniew MV. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016;532:195-200.
- Dong Y, Benveniste EN. Immune function of astrocytes. Glia 2001;36:180-190.
- Endoh M, Maiese K, Wagner J. Expression of the inducible form of nitric oxide synthase by reactive astrocytes after transient global ischemia. Brain Res 1994;651:92-100.
- Donohue PJ, Richards CM, Brown SA, Hanscom HN, Buschman J, Thangada S, Hla T, Williams MS, Winkles JA. TWEAK is an endothelial cell growth and chemotactic factor that also potentiates FGF-2 and VEGF-A mitogenic activity. Arterioscler Thromb Vasc Biol 2003;23:594-600.
- Yepes M, Brown SA, Moore EG, Smith EP, Lawrence DA, Winkles JA. A soluble Fn14-Fc decoy receptor reduces infarct volume in a murine model of cerebral ischemia. Am J Pathol 2005;166:511-520.
- Yilmaz G, Granger DN. Cell adhesion molecules and ischemic stroke. Neurol Res 2008;30:783-793.
- Arvin B, Neville LF, Barone FC, Feuerstein GZ. The role of inflammation and cytokines in brain injury. Neurosci Biobehav Rev 1996;20:445-452.
- Härtl R, Schürer L, Schmid-Schönbein GW, del Zoppo GJ. Experimental antileukocyte interventions in cerebral ischemia. J Cereb Blood Flow Metab 1996;16:1108-1119.
- Chopp M, Li Y, Jiang N, Zhang RL, Prostak J. Antibodies against adhesion molecules reduce apoptosis after transient middle cerebral artery occlusion in rat brain. J Cereb Blood Flow Metab 1996;16:578-584.
- Panés J, Perry M, Granger DN. Leukocyte-endothelial cell adhesion: avenues for therapeutic intervention. Br J Pharmacol 1999;126:537-550.
- Emsley HC, Tyrrell PJ. Inflammation and infection in clinical stroke. J Cereb Blood Flow Metab 2002;22:1399-1419.
- DeGraba TJ. The role of inflammation after acute stroke: utility of pursuing anti-adhesion molecule therapy. Neurology 1998;51:S62-S68.
- Kataoka H, Kim SW, Plesnila N. Leukocyteendothelium interactions during permanent focal cerebral ischemia in mice. J Cereb Blood Flow Metab 2004;24:668-676.
- Gelderblom M, Leypoldt F, Steinbach K, Behrens D, Choe CU, Siler DA, Arumugam TV, Orthey E, Gerloff C, Tolosa E, Magnus T. Temporal and spatial dynamics of cerebral immune cell accumulation in stroke. Stroke 2009;40:1849-1857.
- Garcia-Bonilla L, Moore JM, Racchumi G, Zhou P, Butler JM, Iadecola C, Anrather J. Inducible nitric oxide synthase in neutrophils and endothelium contributes to ischemic brain injury in mice. J Immunol 2014;193:2531-2537.
- Dimasi D, Sun WY, Bonder CS. Neutrophil interactions with the vascular endothelium. Int Immunopharmacol 2013;17:1167-1175.
- Danton GH, Dietrich WD. Inflammatory mechanisms after ischemia and stroke. J Neuropathol Exp Neurol 2003;62:127-136.
- Vemuganti R, Dempsey RJ, Bowen KK. Inhibition of intercellular adhesion molecule-1 protein expression by antisense oligonucleotides is neuroprotective after transient middle cerebral artery occlusion in rat. Stroke 2004;35:179-184.
- Liesz A, Zhou W, Mracskó É, Karcher S, Bauer H, Schwarting S, Sun L, Bruder D, Stegemann S, Cerwenka A, Sommer C, Dalpke AH, Veltkamp R. Inhibition of lymphocyte trafficking shields the brain against deleterious neuroinflammation after stroke. Brain 2011;134:704-720.
- Soriano SG, Coxon A, Wang YF, Frosch MP, Lipton SA, Hickey PR, Mayadas TN, Chan PH. Mice deficient in Mac-1 (CD11b/CD18) are less susceptible to cerebral ischemia/reperfusion injury. Stroke 1999;30:134-139.
- Yenari MA, Kunis D, Sun GH, Onley D, Watson L, Turner S, Whitaker S, Steinberg GK. Hu23F2G, an antibody recognizing the leukocyte CD11/CD18 integrin, reduces injury in a rabbit model of transient focal cerebral ischemia. Exp Neurol 1998;153:223-233.
- Zhang L, Zhang ZG, Zhang RL, Lu M, Krams M, Chopp M. Effects of a selective CD11b/CD18 antagonist and recombinant human tissue plasminogen activator treatment alone and in combination in a rat embolic model of stroke. Stroke 2003;34:1790-1795.
- Kim JS. Cytokines and adhesion molecules in stroke and related diseases. J Neurol Sci 1996;137:69-78.
- Zhang R, Chopp M, Zhang Z, Jiang N, Powers C. The expression of P- and E-selectins in three models of middle cerebral artery occlusion. Brain Res 1998;785:207-214.
- Bargatze RF, Kurk S, Butcher EC, Jutila MA. Neutrophils roll on adherent neutrophils bound to cytokineinduced endothelial cells via L-selectin on the rolling cells. J Exp Med 1994;180:1785-1792.
- Khan M, Jatana M, Elango C, Paintlia AS, Singh AK, Singh I. Cerebrovascular protection by various nitric oxide donors in rats after experimental stroke. Nitric Oxide 2006;15:114-124.
- Huang J, Choudhri TF, Winfree CJ, McTaggart RA, Kiss S, Mocco J, Kim LJ, Protopsaltis TS, Zhang Y, Pinsky DJ, Connolly ES, Feuerstein GZ. Postischemic cerebrovascular E-selectin expression mediates tissue injury in murine stroke. Stroke 2000;31:3047-3053.
- Rossi D, Zlotnik A. The biology of chemokines and their receptors. Annu Rev Immunol 2000;18:217-242.
- Bizzarri C, Beccari AR, Bertini R, Cavicchia MR, Giorgini S, Allegretti M. ELR+ CXC chemokines and their receptors (CXC chemokine receptor 1 and CXC chemokine receptor 2) as new therapeutic targets. Pharmacol Ther 2006;112:139-149.
- Gerard C, Rollins BJ. Chemokines and disease. Nat Immunol 2001;2:108-115.
- Hurn PD, Subramanian S, Parker SM, Afentoulis ME, Kaler LJ, Vandenbark AA, Offner H. T- and B-cell-deficient mice with experimental stroke have reduced lesion size and inflammation. J Cereb Blood Flow Metab 2007;27:1798-1805.
- Brait VH, Rivera J, Broughton BR, Lee S, Drummond GR, Sobey CG. Chemokine-related gene expression in the brain following ischemic stroke: no role for CXCR2 in outcome. Brain Res 2011;1372:169-179.
- Villa P, Triulzi S, Cavalieri B, Di Bitondo R, Bertini R, Barbera S, Bigini P, Mennini T, Gelosa P, Tremoli E, Sironi L, Ghezzi P. The interleukin-8 (IL-8/CXCL8) receptor inhibitor reparixin improves neurological deficits and reduces longterm inflammation in permanent and transient cerebral ischemia in rats. Mol Med 2007;13:125-133.
- Auffray C, Sieweke MH, Geissmann F. Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annu Rev Immunol 2009;27:669-692.
- Ginhoux F, Jung S. Monocytes and macrophages: developmental pathways and tissue homeostasis. Nat Rev Immunol 2014;14:392-404.
- Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K. Development of monocytes, macrophages, and dendritic cells. Science 2010;327:656-661.
- Chu HX, Arumugam TV, Gelderblom M, Magnus T, Drummond GR, Sobey CG. Role of CCR2 in inflammatory conditions of the central nervous system. J Cereb Blood Flow Metab 2014;34:1425-1429.
- Kim JS, Gautam SC, Chopp M, Zaloga C, Jones ML, Ward PA, Welch KM. Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 after focal cerebral ischemia in the rat. J Neuroimmunol 1995;56:127-134.
- Wang X, Feuerstein GZ. Induced expression of adhesion molecules following focal brain ischemia. J Neurotrauma 1995;12:825-832.
- Carlin LM, Stamatiades EG, Auffray C, Hanna RN, Glover L, Vizcay-Barrena G, Hedrick CC, Cook HT, Diebold S, Geissmann F. Nr4a1-dependent Ly6C(low) monocytes monitor endothelial cells and orchestrate their disposal. Cell 2013;153:362-375.
- Dimitrijevic OB, Stamatovic SM, Keep RF, Andjelkovic AV. Absence of the chemokine receptor CCR2 protects against cerebral ischemia/reperfusion injury in mice. Stroke 2007;38:1345-1353.
- Schilling M, Strecker JK, Ringelstein EB, Schäbitz WR, Kiefer R. The role of CC chemokine receptor 2 on microglia activation and blood-borne cell recruitment after transient focal cerebral ischemia in mice. Brain Res 2009;1289:79-84.
- Naert G, Rivest S. A deficiency in CCR2+ monocytes: the hidden side of Alzheimer's disease. J Mol Cell Biol 2013;5:284-293.
- Gliem M, Mausberg AK, Lee JI, Simiantonakis I, van Rooijen N, Hartung HP, Jander S. Macrophages prevent hemorrhagic infarct transformation in murine stroke models. Ann Neurol 2012;71:743-752.
- Li GZ, Zhong D, Yang LM, Sun B, Zhong ZH, Yin YH, Cheng J, Yan BB, Li HL. Expression of interleukin-17 in ischemic brain tissue. Scand J Immunol 2005;62:481-486.
- Stevens SL, Bao J, Hollis J, Lessov NS, Clark WM, Stenzel-Poore MP. The use of flow cytometry to evaluate temporal changes in inflammatory cells following focal cerebral ischemia in mice. Brain Res 2002;932:110-119.
- Becker K, Kindrick D, Relton J, Harlan J, Winn R. Antibody to the alpha4 integrin decreases infarct size in transient focal cerebral ischemia in rats. Stroke 2001;32:206-211.
- Shichita T, Sugiyama Y, Ooboshi H, Sugimori H, Nakagawa R, Takada I, Iwaki T, Okada Y, Iida M, Cua DJ, Iwakura Y, Yoshimura A. Pivotal role of cerebral interleukin-17-producing gammadeltaT cells in the delayed phase of ischemic brain injury. Nat Med 2009;15:946-950.
- Kleinschnitz C, Schwab N, Kraft P, Hagedorn I, Dreykluft A, Schwarz T, Austinat M, Nieswandt B, Wiendl H, Stoll G. Early detrimental T-cell effects in experimental cerebral ischemia are neither related to adaptive immunity nor thrombus formation. Blood 2010;115:3835-3842.
- Saino O, Taguchi A, Nakagomi T, Nakano-Doi A, Kashiwamura S, Doe N, Nakagomi N, Soma T, Yoshikawa H, Stern DM, Okamura H, Mat suyama T. Immunodeficiency reduces neural stem/progenitor cell apoptosis and enhances neurogenesis in the cerebral cortex after stroke. J Neurosci Res 2010;88:2385-2397.
- Dinkel K, Dhabhar FS, Sapolsky RM. Neurotoxic effects of polymorphonuclear granulocytes on hippocampal primary cultures. Proc Natl Acad Sci U S A 2004;101:331-336.
- Liesz A, Suri-Payer E, Veltkamp C, Doerr H, Sommer C, Rivest S, Giese T, Veltkamp R. Regulatory T cells are key cerebroprotective immunomodulators in acute experimental stroke. Nat Med 2009;15:192-199.
- Ren X, Akiyoshi K, Grafe MR, Vandenbark AA, Hurn PD, Herson PS, Offner H. Myelin specific cells infiltrate MCAO lesions and exacerbate stroke severity. Metab Brain Dis 2012;27:7-15.
- Wong CH, Jenne CN, Lee WY, Léger C, Kubes P. Functional innervation of hepatic iNKT cells is immunosuppressive following stroke. Science 2011;334:101-105.
- Harrison DG, Guzik TJ, Goronzy J, Weyand C. Is hypertension an immunologic disease?. Curr Cardiol Rep 2008;10:464-469.
- Barry M, Bleackley RC. Cytotoxic T lymphocytes: all roads lead to death. Nat Rev Immunol 2002;2:401-409.
- Brait VH, Jackman KA, Walduck AK, Selemidis S, Diep H, Mast AE, Guida E, Broughton BR, Drummond GR, Sobey CG. Mechanisms contributing to cerebral infarct size after stroke: gender, reperfusion, T lymphocytes, and Nox2-derived superoxide. J Cereb Blood Flow Metab 2010;30:1306-1317.